
Содержание
Кровеносная система птиц
- Довольно большое сердце. У птиц оно составляет от 1 % массы тела, а у видов с быстрым и маневренным полётом доходит до 2 %. Относительные размеры сердца больше у мелких птиц, чем у крупных.
- Количество ударов сердца в минуту также очень большое. У средних по размеру птиц пульс составляет 200 – 300 ударов в минуту, а в полёте поднимается до 500 ударов. У мелких видов эти показатели существенно выше – в полёте частота работы их сердца достигает 1000 ударов в минуту.
- Эритроцитов и гемоглобина в крови птиц в несколько раз больше, чем в крови пресмыкающихся. Их количество, таким образом, приближается к таковому у млекопитающих. Столь высокая кислородная ёмкость необходима птицам в силу их мощного дыхания, особенно во время полёта. Много в птичьей крови и сахаров.
- Состав крови, её количество и скорость передвижения соответствуют очень быстрому метаболизму в птичьем организме. Лёгкие птиц содержат очень большое количество капилляров. При этом насыщение крови кислородом происходит посредством двойного дыхания: при вдохе только часть кислорода попадает в лёгкие, остальное набирается в так называемые воздушные мешки; при выдохе воздух из воздушных мешков снова проходит через лёгкие и вторично насыщает кровь кислородом.
- Видимо, с «летающим» образом жизни птиц связано и строение их эритроцитов. В отличие от млекопитающих, эритроциты в крови птиц имеют эллиптическую форму и наделены ядрами. Впрочем, стоит отметить, что эритроциты млекопитающих за счёт отсутствия ядер и большинства органелл могут вместить большее количество кислорода.
особенности внутреннего строения кровеносной системы органы птиц
Убедиться в том, что растения дышат, вам помогут несло. проветри Поэтому в зер через окна и д приборов, что Дыханию л них из воздух ца и мешают натные растен Отрицател примеси в во приятий. Вот тов высажин запылённост тополь, лип: ные опыты. Опыт 1. В два одинаковых сосуда нальём воду, в которой раст. ворено небольшое количество минеральных веществ, необходи. мых растению. В каждый сосуд опустим корнями в раствор дро. ростки фасоли, бобов или гороха и закрепим их. Раствор в одном из сосудов ежедневно будем насыщать воздухом с помощью пуль. веризатора. Другой сосуд плотно закроем крышкой так, чтобы в него не проникал воздух. Растения во втором сосуде через некото- рое время погибнут. Сделайте вывод о причине гибели растений. Опыт 2. На дно банки налейте воду и насыпьте до Із её вы- соты прорастающих семян гороха, фасоли или пшеницы. Банку плотно закройте крышкой. В другую банку насыпьте такое же количество сухих семян. Обе банки держите при температуре 20-25 °С. Через сутки опустите в обе банки горящую лучинку. Объясни- те, почему в банке с сухими семенами лучинка будет некоторое время гореть, а в банке с прорастающими семенами лучинка сразу погаснет. Сделайте вывод. Усложните опыт: поставьте одну банку с прорастающими се- менами в холодильник, а другую — в тёплое место. Через один-
Кровь и кровеносная система птиц
 Кровь состоит из красных и белых кровяных телец, кровяных пластинок и плазмы.
Кровь состоит из красных и белых кровяных телец, кровяных пластинок и плазмы.
Красные кровяные тельца (эритроциты) у птиц имеют строение яйцевидной овальной клетки с большим ядром. В этих клетках находится особое красящее белковое вещество (пигмент), содержащее железо; называется это вещество гемоглобином. Гемоглобин обладает способностью легко, но очень непрочно соединяться с кислородом; в таком виде он называется оксигемоглобином; происходит это соединение в легких.
Протекая с током крови по кровеносным сосудам через ткани различных органов, оксигемоглобин отдает кислород тканям, превращаясь снова в просто гемоглобин. Таким образом, красные кровяные тельца разносят кислород по всему организму.
Кроме красных кровяных телец, в крови имеется сравнительно большое количество клеток другого рода, называемых белыми кровяными тельцами, или лейкоцитами. Лейкоциты обладают способностью самостоятельного движения, могут проходить сквозь стенки кровеносных сосудов и проникать во все ткани тела. Они также обладают способностью захватывать посторонние тельца, попавшие в ткани организма, например бактерии, и, захватив, переваривать их. Точно так же лейкоциты уничтожают отмирающие клетки организма, очищая организм от посторонних и вредных веществ. Лейкоциты, кроме того, могут захватывать некоторые питательные вещества, всасывающиеся из пищи, и разносить их по телу. Таким образом, лейкоциты имеют для организма огромное значение. Есть еще в крови третий вид кровяных телец — это так называемые кровяные пластинки, или тромбоциты. Благодаря этим клеткам кровь может свертываться, как только она выходит из кровеносных сосудов, что предохраняет организм от больших кровоизлияний.
Кровь образуется в костном мозгу, селезенке и лимфатических узлах. Кровеносную систему у птиц составляют сердце и кровеносные сосуды.
Сердце — большой кровеносный орган. Оно лежит в грудной области и заключено в околосердечную сумку (сорочку). Верхушка сердца помещается между долями печени, а своим основанием сердце обращено к позвоночному столбу.
Сердце у птиц четырехкамерное, состоит из двух предсердий и двух желудочков, подразделенных на две половины: левую и правую. Таким образом, различают правый желудочек и правое предсердие, левый желудочек и левое предсердие.
Работа сердца заключается в последовательном сокращении и расслаблении сердечной мышцы, что вызывает ритмическое биение сердца. В момент сокращения предсердий и желудочков кровь из них выталкивается, в момент расслабления предсердий и желудочков они наполняются кровью, причем момент расслабления является отдыхом для сердечной мышцы.
Кровеносные сосуды представляют собой замкнутую со всех сторон систему трубок, по которым движется кровь по всему организму. Они подразделяются на артерии, вены и капилляры.
Артериями называются сосуды, которые разносят от сердца по всему организму кровь очищенную, богатую кислородом.
Венами называются сосуды, которые несут к сердцу от отдельных частей тела кровь неочищенную, бедную кислородом. Кровь, находящуюся в артериях, называют артериальной, а в венах — венозной. Эти названия не всегда совпадают с действительностью, например легочная артерия несет венозную кровь, а легочная вена — артериальную.
Капиллярами называются очень мелкие сосуды, которые образуют сеть между конечными разветвлениями артерий и начальными разветвлениями вен. Через капилляры происходит обмен веществ между кровью и тканями. Стенки капилляров, участвуя в обмене, пропускают одни вещества и задерживают другие.
☰
По-сравнению с пресмыкающимися в кровеносной системе птиц произошли два прогрессивных изменения: их сердце стало четырехкамерным, венозная и артериальная кровь нигде не смешиваются. Сердце птиц крупное, сокращается очень часто (сотни раз в минуту), а следовательно кровообращение у них быстрое, кроме того в органы тела поступает чистая артериальная кровь. Все это позволило птицам стать теплокровными животными, температура их тела перестала зависеть от температуры окружающей среды, а значит, они смогли сохранять активность в неблагоприятных температурных условиях (в то время как земноводные и пресмыкающиеся в это время впадают в спячку).
Главным компонентом кровеносной системы позвоночных (в том числе и птиц) является сердце. У птиц сердце состоит из двух предсердий и двух желудочков. Через правую половину сердца (правые предсердие и желудочек) проходит венозная кровь. Через левую половину сердца (левые предсердие и желудочек) проходит артериальная кровь. Следует помнить, что на схемах и рисунках правая половина сердца изображена слева, а левая — справа.
От левого (содержащего артериальную кровь) желудочка отходит одна аорта. У пресмыкающихся отходило две дуги аорты (правая и левая). В кровеносной системе птиц осталось только одна — правая дуга аорты, которая начинает большой круг кровообращения. Выйдя из сердца, аорта разделяется на спинную аорту и сонные артерии. Сонная артерия несет артериальную кровь к голове. Спинная аорта несет кровь к остальным органам тела. От нее отходит множество более мелких артерий.
В капиллярах артериальная кровь, отдав тканям кислород и забрав из них углекислый газ, становится венозной. Далее она собирается в более крупные вены (переднюю полую вену, которая несет кровь от головы, и заднюю полую вену, которая несет кровь от остальных органов тела) и поступает в правое (венозное) предсердие, откуда при его сокращении попадает в правый желудочек сердца. Так завершается большой круг кровообращения.
Малый круг кровообращения у птиц начинается в правом (венозном) желудочке сердца, из которого выходит легочная артерия (хотя называется артерией, но несет венозную кровь). Далее общая легочная артерия ветвится на две легочные артерии (правую и левую), каждая из которых идет в свое легкое. В легких кровь насыщается кислородом и по легочным венам (хотя называются венами, но несут артериальную кровь) возвращается в сердце, в его левое предсердие, откуда попадает в левый желудочек сердца. Так завершается малый круг кровообращения.
Таким образом, кровеносная система птиц имеет два круга кровообращения. Большой круг обеспечивает кровообращение в органах тела, начинается в левом желудочке и заканчивается в правом предсердии. Малый круг обеспечивает кровообращение через легкие, начинается в правом желудочке и заканчивается в левом предсердии.
Следует отметить, что частота сердечных сокращений у птиц в полете обычно возрастает более чем в 2 раза.
Особенности организации птиц (Aves)
Дальнейшая специализация и отбор привели к развитию оперения крыла, обеспечившего возможность сначала планирующего, а затем активного полета. Оперение, покрывающее все тело птиц, вероятно, развивалось не столько как приспособление к полету (обеспечение обтекаемости тела), сколько как приспособление к термоизоляции; высказываются предположения, что оно могло сформироваться еще до выработки способности к полету. У некоторых псевдозухий были удлиненные роговые чешуи с четким продольным гребнем, от которого отходили мелкие поперечные ребрышки. Такие чешуйки, видимо, можно рассматривать как структуру, из которой путем расчленения могло возникнуть птичье перо.
Происхождение и взаимоотношения современных групп (отрядов) птиц остаются, как уже говорилось, за отсутствием достаточных палеонтологических материалов, как правило, очень гипотетическими. Наибольшее внимание уделил этому вопросу М. Фюрбрингер, заметно меньшее — Г. Гадов. Не внесли много нового и бесспорного в эту проблему и современные авторы, в той или иной степени касавшиеся филогении птиц. Поэтому здесь изложено представление о взаимоотношениях отдельных групп птиц преимущественно по Фюрбрингеру, но в ряде случаев, где это нужно, внесены некоторые изменения (в частных случаях эти изменения специально не оговариваются).
Ископаемые остатки веерохвостых птиц обнаружены только в меловом периоде (их возраст примерно 80-90 млн. лет), т. е. они на несколько десятков миллионов лет моложе археоптериксов. По нескольким плохой сохранности остаткам описаны гесперорнисы — очень крупные (длиной более 100 см) птицы, внешне, видимо, несколько похожие на гагар или поганок, совершенно не летавшие (грудина без киля, от передней конечности сохранилось только маленькое тоненькое плечо), но, вероятно, они хорошо плавали и ныряли: челюсти их несли зубы. В меловых же отложениях найдены остатки других зубастых птиц — ихтиорнисов. Они имели хорошо развитый киль грудины, типичное птичье крыло и, видимо, обладали активным полетом. По размерам они были примерно с голубя. Родственные отношения зубастых меловых птиц с другими птицами очень неясны. Возможно, часть этих остатков относится к меловым динозаврам, а не птицам, но часть, несомненно, зубастые птицы. Фюрбрингер сближал гесперорнисов с гагарами и поганками, а их тиорнисов — с чайками, но при этом подчеркивал их очень давнюю обособленность.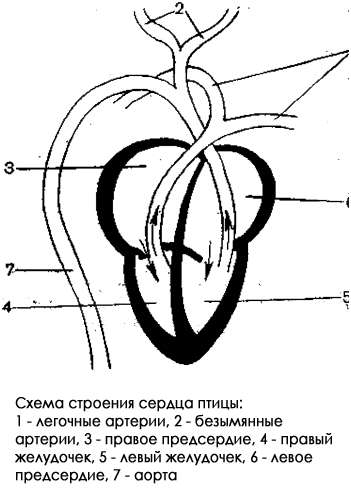
Гадов гесперорнисов выделял в надотряд, показывая этим их обособленное положение, а ихтиорнисы как самостоятельный отряд (тоже обособленный) начинали перечень отрядов килегрудных птиц. Подчеркивая неясность происхождения (да и объем) этих групп, Уэтмор (1960) выделяет их в самостоятельные надотряды.
Из меловых отложений описано еще несколько семейств птиц (по обломкам челюстей и трубчатых костей), но их положение и внешний облик очень неясны: возможно, хотя бы часть этих находок — остатки рептилий. Встреченные в отложениях третичного периода кайнозойской эры (возраст менее 55 млн. лет) остатки птиц с большей или меньшей вероятностью могут быть отнесены к современным отрядам. Во всяком случае они принадлежат к типичным птицам, близким к тем или иным группам современных птиц. Судя по этим материалам, можно предполагать, что интенсивная адаптивная радиация птиц и становление большинства современных отрядов проходило в самом конце мелового периода мезозойской эры — в третичный период кайнозойской эры, т. е. в период примерно 70-40 млн. лет назад.
Гадов всех ныне живущих и ископаемых крупных нелетающих страусоподобных птиц выделял в надотряд бескилевых, подчеркивая этим их сходство и филогенетическое родство (т. е. монофилию), а также их определенную обособленность от остальных групп птиц. Среди килевых птиц наиболее близки к страусоподобным тинаму (скрытохвосты), куриные, журавлеобразные и близкие к ним группы. Фюрбрингер и большинство современных авторов (Уэтмор, Штреземанн и др.) считают страусоподобных птиц сборной полифилетическои группой, разбиваемой на ряд самостоятельных отрядов, начинающих систему, что подразумевает их определенную примитивность и изолированность. Их общие сходные черты — результат конвергенции: крупные размеры, потеря способности к полету, приспособление к быстрому, бегу и т. д.
По Гадову, эволюция остальных птиц шла в двух направлениях (две ветви), каждое из которых в свою очередь разделяется на 2 группы родственных отрядов. Поганко-гагарообразные (ихтиорнисы, поганки, гагары, пингвины и трубконосые) родственны группе Pelargomorphae (аистообразные, гусеобразные, хищники), а близкая к страусоподобным группа курообразных (тинаму, куриные, журавлеобразные, куликообразные) дала начало ракшеобразным (кукушки, ракши, воробьиные).
Эволюционные представления Фюрбрингера в общих чертах близки к этой схеме Гадова (об иных взглядах на страусоподобных говорилось выше), но он излагает свои представления с большей степенью детализации. Крупный ствол птиц — отряд Pelargornithes — делится на ряд ветвей: гусеобразных, поганкообразных (включая поганок, гагар, гесперорнисов); от основания этих ветвей отходит еще одна мощная ветвь, которая, разделяясь, дает начало веслоногим, дневным хищникам и группам подотряда аистообразных.
Отряд паламедей Palamedeiformes занимает промежуточное положение между нанду и отрядом Pelargornithes. Промежуточные отряды пингвинов Aptenodytiformes и трубконосых Procellariiformes довольно обособлены; возможно, они имеют давние слабые связи друг с другом и, еще более отдаленные, с отрядами Pelargornithes и куликообразных Charadriornithes. Ихтиорнисы Ichthyornithiformes — тоже промежуточный отряд, но с несколько более выраженными давними связями с куликообразными. Отряд куликообразных Charadriornithes очень давними связями видимо может быть приближен к отряду ракшеобразных. Основные ветвления ствола куликообразных — подотряд Laro-Limicolae — кулики, чайки и чистики; к ним довольно близки зобатые бегунки и дрофы (переходной группой между куликами и дрофами служат авдотки). От основания ствола куликообразных обособились еще две крупные ветви: журавлеобразные Gruiformes и пастушковые Ralliformes. Журавлеобразные разделяются на несколько ветвей: собственно журавлиных (журавли, арамиды, трубачи, кариамы) и занимающих несколько обособленное положение солнечных цапель и кагу. Пастушковые подразделяются на родственные ветви: пастушков, лапчатоногих и стоящих несколько обособленно трехперсток и мадагаскарских пастушков.
Один из древних стволов родословного древа птиц — отряд курообразных Alectorornithis. Наиболее примитивная ветвь — подотряд бескрылых, или киви Apterygiformes, включающий киви и моа; обе группы разошлись почти сразу после своего обособления. Общим происхождением с предыдущей ветвью, видимо, связаны тинаму (или скрытохвосты) Crypturiformes, в свою очередь через предков близкие к подотряду куриных Galliformes (практически в современном объеме). Отряд курообразных очень давними родственными связями, вероятно, более или менее близок к пастушковым Ralliformes, а через них и ко всему отряду куликообразных.
Промежуточные отряды голубеобразных Columbiformes (включающий рябков и голубей) и попугаев Psittaciformes занимают в системе обособленное положение. Вероятно, через очень давние отношения они как-то связаны с обширным отрядом ракшеобразных. По представлениям Гадова, голуби и рябки в ранге подотрядов входят в отряд куликообразных, а попугаи — тоже как подотряд — в отряд кукушкообразных. По сути дела сходной точки зрения придерживается и Уэтмор, ставящий отряд голубеобразных сразу же за отрядом куликообразных, а отряд попугаев — рядом с отрядом кукушек.
Обширный и разнообразный отряд Coracornithes своими истоками связан с предками отряда куликообразных. Основная и наиболее мощная ветвь — подотряд дятлов и воробьиных Pico — Passeriformes, которая затем разделяется на отдельные группы собственно воробьиных и дятловых. От основания этой ветви обособились Macrochires (стрижи, колибри) и птицы мыши Colii. Еще раньше от этой ветви обособляются трогоны Тгоgones. От самого основания ветви дятлово-воробьиных птиц обособляются и три других подотряда ракшеобразных. Подотряд кукушкообразных Coccygiformes отделяет ветвь якамар Galbulae (якамары и пуховки, а по Уэтмору — это подотряд отряда дятлов), а затем разделяется на кукушек Cuculidae и бананоедов Musophagidae. Подотряд ракшеобразных Coraciiformes разделяется на собственно ракш Coraciae и на вторую ветвь, вскоре делящуюся на сов Striges и козодоев Caprimulgi. Подотряд зимородковых Haleyoniformes (он, видимо, ближе к Pico — Passeriformes, чем к другим подотрядам), отделяя в самом начале боковую веточку тоди Todi (кроме тоди Фюрбрингер сюда включает и момотов), разделяется на 3 родственные группы: зимородковых Halcyones (Alcedinidae), удодов Bucerotes (удоды, птицы-носороги) и щурок Meropes.
Сопоставление филогенетических представлений Фюрбрингера со взглядами Гадова и Уэтмора (очередность и порядок перечисления отрядов) показывает значительное их совпадение; резкие отличия в положении и взаимоотношении отдельных групп указаны выше.
В последние годы попытку графического изображения филогенетических взаимоотношений между отдельными группами птиц предприняли немецкие орнитологи Берндт и Майзе в капитальной трехтомной сводке «Естественная история птиц». Подкласс древних ящерохвостых птиц Palaeornithes дал начало подклассу новых веерохвостых птиц Neornithes, который включает 24 отряда, подразделяемых авторами обычно сразу на семейства; таксономические категории ранга подотряда и надсемейства используются, как правило, только в отряде воробьиных. Взаимоотношения между отрядами птиц, по Берндту и Майзе, выглядят следующим образом.
От древних птиц эволюция шла по двум направлениям (ветвям): ветви (надотряд) наземных и водных птиц Geornithes и ветви (надотряд) древесных птиц Dendrornithes. От самого начала ветви наземных и водных птиц общим стволом отделились, позже разделившись, тинаму Crypturi и бескилевые птицы Ratitae, включая всех ныне живущих страусоподобных, в том числе и киви, и ископаемых моа и эпиорнисов. Далее обособляются куриные птицы Galli (в полном объеме отряда Уэтмора) и на этом же уровне, но самостоятельной ветвью — гагарообразные Pygopodes (включающие ископаемых меловых зубастых птиц гесперорнисов, гагар и поганок). Все остальные отряды этой ветви (надотряда) имеют общее происхождение — представляют собой как бы мутовку стволов, отходящих от одного участка. Относительно тесно связаны друг с другом и несколько противопоставляются другим группам отряды, журавлеобразных (пастушки, мадагаскарские пастушки, солнечные цапли в лапчатоноги, кагу, настоящие журавли, арамы, трубачи, дрофы, кариамы, трехперстки; из ископаемых — диатримы и форораки Phororhacidae) и болотно- и чайкообразных птиц Limicolae — Lari (яканы, различные группы собственно куликов, чайки чистики и — из ископаемых — зубастые меловые птицы — ихтиорнисы). Далее от общего основания мутовки (т. е. имея общих предков) последовательно обособляются отряды гусеобразных Anseres (паламедеи и собственно гусеобразные), фламинго Phoenicopteri, голенастые Gressores, дневные хищники Accipitres и веслоногие Stegaribpodes. От форм, близких к предкам веслоногих, отделяется ветвь, которая затем разделяется на отряды трубконосых Tubinares и пингвинов Sphenisci.
Развитие второй ветви (надотряда) древесных птиц Dendrornithes начинается от каких-то предковых форм, видимо, более древних, чем предки первой ветви (первого надотряда). Сначала общим стволом обособляются 3 отряда: кукушкообразных Cuculi, голубей Columbae (голубки, рябки и дронты) и попугаев Psittaci. Следующая обособившаяся ветвь разделяется на 2 ответвления, каждое из которых образует по 2 отряда. Одно из них дает начало совам Striges и козодоям Caprimulgi, другое — трогонам Trogones и ракшеобразным Coracii. Дальше от общего ствола также общей ветвью обособляются отряды птиц-мышей Colii и стрижеобразных Macrochires (стрижи и колибри), самостоятельной ветвью отходит отряд дятлов Pici (в объеме Уэтмора), а основной ствол ветвится на многочисленные подразделения отряда воробьиных Passeres. Помимо этого, авторы дают гипотетические схемы, в которых пытаются представить филогенетические взаимоотношения в пределах нескольких отдельных отрядов (дневных хищников, веслоногих, гагарообразных и воробьиных).
Филогенетические схемы Берндта и Майзе ясны, наглядны и легко запоминаются. Однако они практически совсем не аргументированы, не обоснованы материалами и, видимо, в значительно меньшей степени отражают действительные родственные (филогенетические) отношения между отдельными группами птиц, чем соответствующие схемы Фюрбрингера. Теоретически маловероятно, что уже на начальных стадиях эволюции класса четко определилась последующая биотопическая специализация птиц, что выражается в изложенной схеме деления подкласса «новых птиц» только на 2 обособленных надотряда и в почти линейном, последовательном обособлении отрядов. Оценивая по современным формам присущий птицам характер эволюции, выражающийся в широкой адаптивной радиации в пределах отдельных групп (о чем кратко упоминалось выше), кажется более логичным предполагать, что подобная же широкая адаптивная радиация была характерна и для начальных фаз эволюции класса. Если это так, то можно говорить, что филогенетические схемы Фюрбрингера хотя бы внешне действительно отражают преобладание такого типа эволюции.
Сейчас еще рано говорить о вполне сложившейся системе класса. Однако системы Фюрбрингера, Гадова и Уэтмора, видимо, могут рассматриваться как хорошая основа для построения естественной системы. Их материалы дают и самое общее представление о филогенетических взаимоотношениях большинства групп. Для улучшения и уточнения системы класса птиц необходимы дальнейшие разнообразные исследования на всех таксономических уровнях (от подвидовых группировок до отрядов и надотрядов): морфологические, экологические, этологические, генетические. Возможно, в будущем ценные материалы даст применение биохимических и физиологических показателей; использование современных методик пока существенного вклада в таксономию птиц не внесло. Хочется надеяться, что увеличится и количество палеонтологических материалов, которые позволят более обоснованно обсуждать эволюцию птиц в целом и отдельных групп (отрядов, семейств).
Особенности кровообращения у птиц
ü Редукция левой дуги аорты
ü Почти полная редукция воротной системы почек (началась у пресмыкающихся)
ü Крупное сердце и высокая интенсивность работы (у птицы массой 500г в покое частота сердечных сокращений 250 уд/мин, в полете 500 уд/мин)
ü Высокое кровяное давление – 120 200 мм рт ст. (у рептилій 30-50 мм рт ст., у млекопитающих 70-160 мм тр ст.)
ü Редукция брюшной вены (замещена копчиково-брыжеечной)
ü Только часть крови движется по почечным капиллярам, остальная – по продолжениям воротных вен почек общим подвздошным венам.
ü Сердце четырехкамерное, два круга кровообращения => полное разделение артериальной и венозной крови в сосудах => уровень обменных процессов повысился => птицы стали гомойотермными животными
Нервная система.
Спинной мозг сегментирован. Длина спинного мозга короче позвоночного столба.Усиливается контроль головного мозга над рефлексами спинного, улучшается иннервация конечностей.
Головной мозг зауропсидного типа – высший центр интеграции – передний мозг,
 отходит 12 пар черепно-мозговых нервов
отходит 12 пар черепно-мозговых нервов
 конечный мозг: укрупнениеполушарий, обонятельные доли очень малы, свод мозга
конечный мозг: укрупнениеполушарий, обонятельные доли очень малы, свод мозга
первичный, с небольшими участками серого вещества, основная масса которого
приходится на полосатые тела – разрастания дна переднего мозга (основной
интегрирующий центр нервной системы),особенно хорошо развиты базальные ядра полосатого тела (ассоциативные реакции)
промежуточный мозг: небольшой, эпифиз развит слабо, гипофиз крупный из трех долей,
таламус, гипоталамус + гипофиз => координация работы эндокринной системы,
средний мозг: крупные зрительные доли отодвинуты в стороны => высший анализ
сигналов от органов зрения
мозжечок развит очень хорошо => сложнейшие высоко координированные движения
 продолговатый мозг – образует изгиб в вертикальной плоскости, характерный для всех
продолговатый мозг – образует изгиб в вертикальной плоскости, характерный для всех
амниот, является центром безусловных рефлексов и вегетативных функций.
Органы чувств:
 глаза — выпуклая роговица, двояковыпуклый хрусталик, двойная аккомодация
глаза — выпуклая роговица, двояковыпуклый хрусталик, двойная аккомодация
(изменение кривизны хрусталика и перемещение его за счет сокращения особых
кольцевых мышц, окружающих склеру), в стекловидное тело вдается гребень (богат
сосудами, крупнее и сложнее, чем у рептилий), в склере – кольцо из уплощенных
тонких костей (прочность крупных глаз), подвижные веки и мигательная перепонка.
Сетчатка: палочки и колбочки в разном сочетании позволяют различать детали
объекта при низкой освещенности (у дневных хищников 2 желтых пятна – питаются
подвижной добычей). Очень подвижный зрачок (предотвращает засветку сетчатки при
быстрых поворотах в полете). Зрение бинокулярное (зрительные оси обоих глаз
совпадают) и монокулярное (у каждого глаза своя ось →охват бокового пространства).
Цветовое зрение (яркое оперение);
органы слуха – внутреннее ухо (улитка обособлена от преддверия) для звуковосприятия и
среднее ухо (барабанная перепонка на дне воронки – зачатка слухового прохода,
может окружаться кожной складкой и перьями, стремечко, евстахиевы трубы)
позволяет улавливать, концентрировать и направлять звуки к барабанной перепонке.
Острый слух, способность к акустическому анализу → способность издавать звуки,
несущие биологически важную информацию.
органы равновесия — 3 полукружные каналы, овальный и круглый мешочек, в эндолимфе
содержатся отолиты, которые давят на чувствительные клетки, возбуждая их;
органы обоняния –разрастаются носовые раковины и S обонятельного эпителия (у
падальщиков, куликов, воробьиных обоняние используется при поисках пищи, у киви
обоняние — ведущий рецептор, а глаза малы);
органы вкуса – вкусовые почки на слизистой рта, языка; птицы распознают сладкое, горькое,
соленое.
ü ↓ интенсивность фильтрации => ↓потери Н2О.
ü Конечный продукт – мочевая кислота (легко выпадает из раствора кристаллами)
=> редукция мочевого пузыря.
ü Мочевая кислота поступает в просвет почечных канальцев не фильтрацией, а путем
секреции железистыми клетками стенок канальцев.
ü В клоаке происходит дополнительное всасывание воды.
Объем мочи невелик, моча концентрированная. Количество мочи настолько мало, что Выделительная системапредставлена тазовыми (1-2% массы тела) почками,
морфофункциональными единицами которых являются нефроны рептильного типа (проксимальные канальцы → дистальные канальцы) и маммального типа (проксимальный каналец → петля Генле → дистальный каналец). Конечный продукт обмена мочевая кислота (нерастворима, много воды для выведения не надо), поэтому многие птицы, довольствуются водой, содержащейся в пище. Мочевого пузыря нет.
тазовые почки компактные тела, расположенные у спинной стенки тазовой области,
состоят из коркового слоя (тельца + канальца) и мозгового слоя (петля Генле и
собирательная трубочка),
мочеточники, железистые клетки эпителия слизь, содержащую муцин для защиты от
повреждений стенок кристаллами солей мочевой кислоты,
клоака – дополнительное всасывание Н2О
Нефрон — структурная и функциональная единица почки
клубочек капилляров — приносящий и выносящий сосуды с артериальной кровью, размеры
невелики => ↓ фильтрации и ↓потери воды;
боуменова капсула – слепые расширения, окружающие клубочек капилляров, внутренняя
стенка которого образована плоским эпителием (ультрафильтрация);
проксимальный каналец – выстлан железистым эпителием, оплетен кровеносными
сосудами; происходит реабсорбция Н2О, сахаров, витаминов, ионов Na;
петля Генле – пассивно выделяется Н2О, удаляются соли => объем мочи↓, моча становится
концентрированной (минимальная потеря Н2О);
дистальный каналец – теряется Н2О, мочевая кислота → в просвет почечных
канальцев путем секреции специальными железистыми клетками стенок
канальцев, оплетенных капиллярами воротной системы почек, открываются в.
собирательные трубочки