
Классификация
В старых классификациях протистов в состав типа Саркомастигофоры включали два подтипа — Саркодовые (Sarcodina) и Жгутиконосцы (Mastigophora) (оба они также представляют собой полифилетические группы, как и многие классы из приведённой ниже классификации).
Подтип Саркодовые
- Класс Саркодовые (Sarcodina)
- Подкласс Корненожки (Rhizopoda)
- Отряд Амёбы (Amoebina)
- Род Амёбы, или Голые амёбы (Amoebina)
- Вид Амёба протей (Amoeba proteus)
- Род Амёбы, или Голые амёбы (Amoebina)
- Отряд Раковинные амёбы (Testacea)
- Отряд Фораминиферы (Foraminifera)
- Род Globobulimina
- Род Hyperammina
- Вид Hyperammina elongata
- Вид Hyperammina elongata
- Вид Hyperammina friabilis Brady
- Вид Hyperammina levigata Wright
- Род Rhabdammina
- Род Astrorhiza
- Вид Astrorhiza limicola
- Род Myxotheca
- Вид Myxotheca arenilega
- Отряд Амёбы (Amoebina)
- Подкласс Лучевики, или Радиолярии (Radiolaria)
- Отряд Акантарии (Acantharia)
- Род Achanthometra
- Вид Achanthometra elastica
- Род Achanthometra
- Отряд Спумеллярии (Spumellaria)
- Род Thalassicola
- Вид Thalassicola muleata
- Род Chromyodrymus
- Вид Chromyodrymus abiatanus
- Род Thalassicola
- Отряд Назеллярии (Nasselaria)
- Род Medusetta
- Вид Medusetta craspedota
- Род Medusetta
- Отряд Феодарии (Phaeodaria)
- Род Aulacantha
- Вид Aulacantha scolymantha
- Род Aulacantha
- Отряд Стихолонхеи (Sticholonchea)
- Род Sticholonche
- Отряд Акантарии (Acantharia)
- Подкласс Солнечники, или Гелиозои (Heliozoa)
- Отряд Актиносфериды (Actinosphaerium)
- Род Actinosphaerium
- Вид Actinosphaerium eichhorni
- Род Actinosphaerium
- Отряд Актиносфериды (Actinosphaerium)
- Подкласс Корненожки (Rhizopoda)
Подтип Жгутиковые
- Класс Жгутиконосцы, или Жгутиковые (Mastigophora)
- Подкласс Растительные жгутиконосцы (Phytomastigina)
- Отряд Хризомонады (Chrysomonadina)
- Род Dinobryon
- Род Synura
- Отряд Динофлагелляты (Dinoflagellata, или Peridinea)
- Род Noctiluca
- Отряд Эвгленовые (Euglenoidea)
- Род Euglena
- Вид Эвглена зелёная
- Род Euglena
- Отряд Вольвоксовые (Volvocidae)
- Род Вольвокс (Volvox)
- Вид Вольвокс (Volvox)
- Род Вольвокс (Volvox)
- Отряд Фитомонады (Phytomonadina)
- Род Chlamydomonas
- Отряд Хризомонады (Chrysomonadina)
- Подкласс Животные жгутиконосцы (Zoomastigina)
- Отряд Воротничковые жгутиконосцы (Choanoflagellata)
- Род Codosiga
- Род Sphaeroeca
- Отряд Ризомастигины (Rhizomastigina)
- Отряд Кинетопластиды (Kinetoplastida)
- Род Bodo
- Род Трипаносомы (Trypanosoma)
- Вид Trypanosoma rhodesiense
- Вид Trypanosoma evansi
- Вид Trypanosoma brucei
- Вид Trypanosoma equiperdum
- Род Лейшмании (Leishmania)
- Вид Leishmania donovani
- Вид Leishmania tropica
- Отряд Полимастигины (Polymastigina)
- Род Трихомонады (Trichomonas)
- Вид Trichomonas hominis
- Вид Trichomonas vaginalis
- Род Лямблии (Lamblia)
- Вид Lamblia intestinalis
- Род Трихомонады (Trichomonas)
- Отряд Гипермастигины (Hypermastigina)
- Род Lophomonas
- Отряд Опалины (Opalina)
- Род Опалины (Opalina)
- Вид Опалина лягушачья (Opalina ranarum)
- Род Опалины (Opalina)
- Отряд Воротничковые жгутиконосцы (Choanoflagellata)
- Подкласс Растительные жгутиконосцы (Phytomastigina)
В период использования этой системы зоологами ботаники (более обоснованно) рассматривали разные «отряды» растительных жгутиконосцев как представителей различных отделов водорослей. Яркий пример — эвгленовые, которые в ботанической систематике имеют ранг отдела Euglenophyta.
В некоторых вузовских и большинстве школьных учебников данная система сохраняется до сих пор. В научной литературе, посвящённой протистам, эта система не используется с 90-х годов XX века. В некоторых из современных классификаций протистов ранг таксонов не указывается — видимо, в частности для того, чтобы его было удобнее изменять в будущем.
Характеристика жгутиковых
Жгутиковые обширный класс включающий более 6000 видов разнообразных простейших.
Тело жгутиконосцев имеет весьма разнообразную, чаще овальную, шаровидную или веретеновидную форму. Органеллами движения служат цитоплазматические нитевидные жгутики. Чаще всего их один или два, но может быть четыре и больше. Жгутики обычно помещаются на переднем конце тела и направлены вперед, по движению животного. Однако у некоторых форм при наличии двух и более жгутиков один из них может загибаться вдоль тела назад и иметь значение руля. Этот жгутик обычно соединен с поверхностью тела тончайшей плазматической пленкой, * укрепляя таким образом колеблющуюся, или волнообразную, перепонку. Движения направленных вперед жгутиков довольно сложны и разнообразны. Жгутик совершает вращательное движения, описывая фигуру в форме конуса. В то же время он сам волнообразно изгибается. Движение жгутика вызывает вращательное движение тела простейшего, которое как бы ввинчивается в воду. У некоторых жгутиконосцев (отряд Rhizomastigida), кроме постоянных жгутиков, ршеются псевдоподии. Эти формы указывают на близость классов жгутиковых и саркодовых.
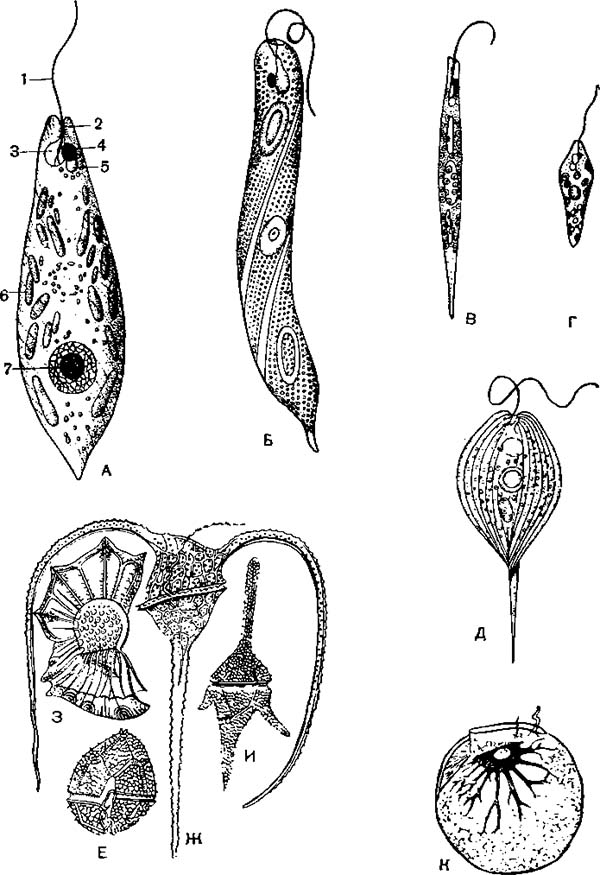 Различные жгутиконосцы:
Различные жгутиконосцы:
А—Д—овгленовые: А — зеленая эвглена (Euglena viridis); 1, 2 — жгут; 3 — резервуар сократительной вакуоли; 4 — стигма; 5 — сократительная вакуоля; 6 — хроматофоры; 7—ядро; Б — Е. oxyuris; В — Е. acus; Г — Е. gracilis; Д — Phacus longicauda; Е—К — панцирные: Е —* Peridinium; Ж — Ceratium tripos; 3 — Ornithocercus; И — Ceratium hirudinella; К -> ночесветка
(Noctiluca miliaris).
Тело жгутиковых состоит из цитоплазмы и обычно одного пузыревидного ядра. Цитоплазма разделяется на два слоя: наружный, более плотный, гомогенный и прозрачный — эктоплазму и внутренний, более жидкий и зернистый — эндоплазму.
Тело большинства покрыто довольно плотной эластичной оболочкой — пелликулой, образующейся за счет уплотнения эктоплазмы. Пелликула позволяет животному временно изменять форму тела, или метаболировать. У некоторых из них тело покрыто твердой оболочкой из клетчатки, которая образует своеобразный панцирь.
Питание
Типы питания простейших наиболее разнообразны в классе жгутиковых. Только среди них встречаются формы, способные, подобно растениям, к фотосинтезу, т. е. к синтезу различных углеводов из ранее поглощенных углекислого газа и воды, происходящему на солнечном свету. Белки их тела строятся из синтезированных углеводов и поглощенных из окружающей среды неорганических солей, в том числе азотистых. Таких жгутиконосцев называют аутотрофными, а их способ питания голофитным или растительным.
К жгутиконосцам с голофитным типом питания относятся все организмы, имеющие хлорофилл. Они питаются так же, как и все зеленые растения. Органеллами питания у таких жгутиконосцев служат окрашенные тельца — хроматофоры различной формы и величины (рис. А), содержащие хлорофилл и другие пигменты (желтый, красный). Благодаря маскирующему влиянию этих пигментов многие жгутиконосцы не имеют зеленой окраски, несмотря на наличие хлорофилла. Жгутиконосцы с голофитным питанием часто имеют уже упомянутую выше клетчатковую оболочку. Это еще более подчеркивает их принадлежность к растениям.
Другие жгутиконосцы питаются органическими продуктами распада более сложных веществ, которые осмотическим путем поступают внутрь их тела. Такой тип питания называют сапрофитным, а организмы, питающиеся таким способом,— сапрофитами. Сапрофиты уже не ауто- трофные (самопитающиеся) организмы, а гетеротрофные, т. е. питающиеся готовым органическим веществом. жгутиконосцы — сапрофиты живут в загрязненных водоемах, богатых разлагающимися органическими веществами. Потребляя их, они тем самым участвуют в очистке воды подобных водоемов.
Многие паразитические жгутиконосцы и другие простейшие питаются также сапрофитным способом, всасывая всей поверхностью тела сыворотку крови, тканевые жидкости или (кишечные паразиты) продукты переваривания пищи их хозяина.
Наконец, третью группу составляют жгутиконосцы, питающиеся, как типичные животные, твердой пищей: бактериями, мелкими водорослями и т. п. Они имеют специальные органеллы питания: рот, глотку и пищеварительные вакуоли; последние образуются в плазме по мере заглаты-вания пищи. Такой тип питания называют голозойным или анимальным.
Особое положение по способу питания занимают некоторые эвглены, в частности зеленая эвглена (Euglena viridis) (рис. А). У зеленой эвглены имеются хроматофоры, содержащие хлорофилл. На свету она питается как типичный зеленый растительный организм. Но, помещенная в темноту, эвглена обесцвечивается и переходит к сапрофитному питанию. По-видимому, и на свету эвглена обладает смешанным способом питания, т. е. является миксотрофным организмом.
Голофитные жгутиконосцые обычно имеют красное или бурое пятнышко — глазок, или стигму,— расположенное на переднем конце тела у основания жгутика (рис. А). Это — светочувствительная органелла, которая обусловливает способность зеленых жгутиковых воспринимать световые раздражения. Зеленые жгутиконосцы обнаруживают положительную реакцию на свет, т. е. обычно передвигаются по направлению к источнику света.
У многих пресноводных имеются специальные органеллы выделения — сократительные вакуоли, при помощи которых из клетки удаляется избыток воды, а также жидкие продукты диссимиляции. Сократительная вакуоля у эвглены (рис. А) помещается в передней части тела. Сократительная в’акуоля периодически наполняется жид-
кпм содержимым, которое при сжатии вакуоли выталкивается в особый: пузырек — резервуар (рис. Л), а затем через особый канал, иногда неправильно называемый «глоткой», выделяется наружу. Сократительные* вакуоли, как правило, отсутствуют у морских жгутиковых. Кроме описан-ных органелл, в цитоплазме могут быть различные включения: зерна крахмала (у зеленых жгутиковых), зерна парамила — углевода, близкого по составу к крахмалу (у эвглены и др.), капельки жира и т. п.
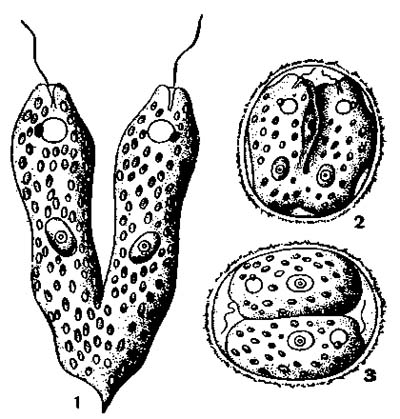 Размножение Эвглены зеленой
Размножение Эвглены зеленой
1 — продольное деление; 2 и 3 — деление в цисте.
Инцистирование
Для жгутиконосцев, так же как и для многих других простейших, характерно образование покоящихся стадий путем инцистирования. Образование цист может иметь различное значение. Инцистирование наблюдается при наступлении неблагоприятных условий: цисты выносят резкие изменения температуры и недостаток влажности. В других случаях инцистирование может быть связано с размножением или представлять собой приспособление, обеспечивающее распространение животных.
Размножение
Для большинства жгутиконосцев известно только бесполое размножение. Половой процесс изучен лишь у зеленых колониальных форм из семейства вольвоксовых (отряд-Phytomonadida).
Бесполое размножение происходит обычно путем деления надвое. Оно начинается с митотического деления ядра, за которым следует продольное деление тела, начинающееся с его переднего конца. При этом делятся также базальные тельца; жгутики или отбрасываются, или переходят к одной из дочерних особей, а у другой развиваются заново из базального тельца. При делении жгутиковых, имеющих хроматофоры, последние также делятся и распределяются по дочерним клеткам. У некоторых жгутиковых делению предшествует инцистирование, при этом деление происходит уже под оболочкой цисты (рис. 2 и 3).
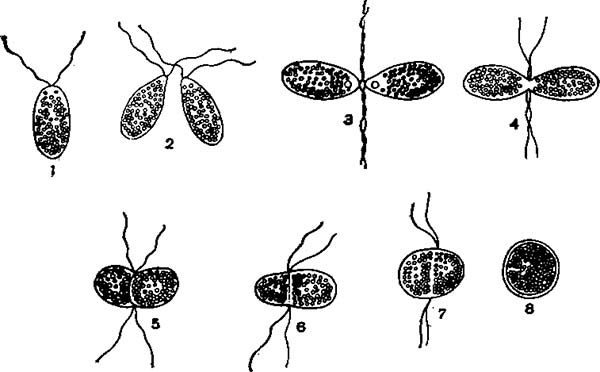 Изогамная копуляция у Polytoma uvella:
Изогамная копуляция у Polytoma uvella:
U — гамета; 2—7 — последовательные стадии копуляции гамет; 8 — инцистированная зигота.
У колониальных жгутиковых из вольвоксовых при бесполом размножении отдельная клетка колонии делится несколько раз (по числу клеток составляющих колонию), и таким образом возникают дочерние колонии.
Половое размножение у разных жгутиковых происходит различно. В простейшем случае происходит копуляция двух внешне одинаковых ш> величине половых особей — гамет, например у Polytoma и некоторых видов хламидомонад. Такую копуляцию одинаковых гамет, или изогамет, называют изогамной копуляцией. У других жгутиковых гаметы отличаются размерами. Одни из них более крупные — макрогаметы, другие более мелкие — микрогаметы. Макро- и микрогаметы снабжены жгутиками. Макро- и микрогаметы копулируют друг с другом. Копуляцию неравных гамет, или анизогамет, называют анизогамной.
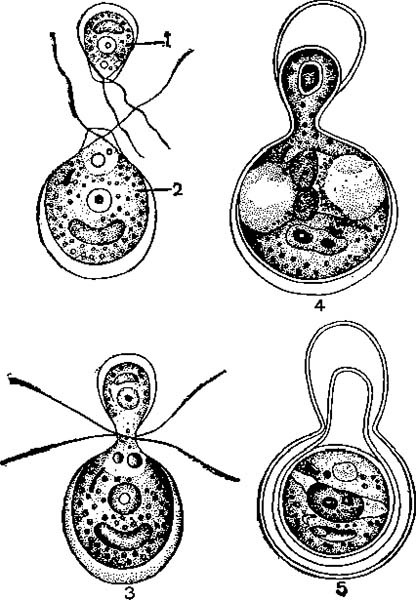 Анизогамная копуляция у Chlamydomonas braunii:
Анизогамная копуляция у Chlamydomonas braunii:
l — микрогамета; 2 — макрогамета; 3—5— последовательные стадии копуляции гамет.
Формирование анизогамет можно считать первым этапом дифференциации пола. У колоний Volvox, образованных огромным числом особей (до 10—20 тыс.), гаметы могут формироваться только из отдельных генеративных особей. Большинство особей колонии вегетативпые, не способные участвовать в половом размножении. В колонии Volvox только 25—30 клеток (особей) может превращаться в крупные неподвижные макрогаметы и лишь 5—10 клеток—образовывать микрогахметы. При формировании макрогамет деления клеток не происходит, а при образовании микрогамет происходит ряд делений, в результате которых возникают 256 мельчайших микрогамет, каждая с парой жгутиков. Микрогаметы покидают колонию и выходят в воду. Затем они проникают в другие колонии, где копулируют с макрогаметами.
Макрогаметы жгутиками не обладают. Их можно сравнить с яйцевыми клетками многокле-точных, а микрогаметы — со сперматозоидами. Такую копуляцию Volvox называют о о г а- м и е й. После оогамной копуляции образуется оплодотворенная макрогамета — зигота. Последняя выделяет плотную оболочку, а все вегетативные особи колонии погибают. Затем зигота начинает многократно делиться и дает начало новой колонии Volvox.
У Volvox globator микро- и макрогаметы развиваются в одной и той же гермафродитной колонии, тогда как у другого вида — V. aureus — колонии раздельнополы: одни из них (женские) образуют только макрогаметы, другие (мужские) — только микрогаметы.
Таким образом, у различных представителей жгутиковых наблюдаются как бы различные ступени эволюции полового процесса — от изогам- ной копуляции до настоящей оогамии, что представляет интерес для понимания происхождения полового процесса у многоклеточных.
Классификация
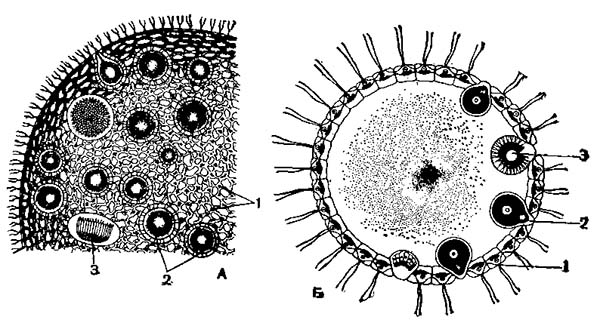 Вольвокс (Volvox globator)
Вольвокс (Volvox globator)
А — часть полонии; Б — схематический разрез вольвокса; 1 — клетки колонии; 2 — макрогаметы; 3 — формирование микрогамет.
Класс жгутиковых (Mastigophora, или Flagellata) состоит из большого числа (13—14) отрядов. Главнейшие из них следующие: J. Эвглеио- вые (Euglenida); 2. Панцирные (Dinoflagellata); 3. Фитомонадовые (Pliy- tomonadida); 4. Корнежгутиковые (Rhizomastigida); 5) Протомонадовыо (Protomonadida); 6. Многожгутиковые (Polymastigida); 7. Гипермастиги- новые (Hypermastigida); 8. Опалиновые (Opalinida).
Первые три отряда (эвгленовые, панцирные, фитомонадовые) входят в состав подкласса фитомастигины (Phytomastigina) — аутотрофнтлх растительных жгутиконосцев. Следующие четыре отряда (корнежгутиковые, протомонадовые, многожгутиковые, гипермастигиновые) составляют подкласс зоомастигины (Zoomastigina) — типичных гетеротрофных организмов. Последний отряд (опалиновые) многие ученые выделяют в ранг самостоятельного подкласса Opalinina.
Классификация
Жизненный цикл паразитов, его экологическое и медицинское значение
Жизненный цикл паразита – совокупность всех стадий онтогенеза паразита и пути передачи его от одного хозяина к другому. Носит циклический характер. Различают:
Простой – развитие, включающее две формы — вегетативную и цисту (амебы).
Сложные — развитие с превращением и многими стадиями.
Свободноживущие организмы довольно легко преодолевают проблемы размножения и расселения, что значительно усложнено у паразитов. Поэтому большинство паразитов нередко развиваются со сложным метаморфозом, включающим много личиночных стадий, обитающих в разных средах и выполняющих разные функции: расселения, активного роста, пассивного ожидания попадания в другую среду обитания и иногда даже размножения.
Личинки могут вести как свободный, так и паразитический образ жизни. Хозяин, в котором обитают личинки паразита, носит название промежуточного. Значение промежуточных хозяев в циклах развития паразитов очень велико: они являются источниками заражения окончательных хозяев, часто выполняют расселительные функции, а иногда обеспечивают выживание популяций паразита в случае временного исчезновения окончательных хозяев.
3.21 Тип Простейшие. Классификация (по латыни). Характерные черты организации. Медицинское значение класса инфузорий
К типу простейших (Protozoa) относится ряд патогенных для человека форм, поражающих отдельные ткани и органы и вызывающих заболевания различной тяжести, в том числе и с летальным (смертельным) исходом.
Морфофизиологическая характеристика. По строению соответствуют отдельной клетке многоклеточных (отсюда название «одноклеточные»), по функциям — целому самостоятельному организму. Организм простейших, передвигается, захватывает пищу, размножается, защищается от врагов, т. е. обладает всеми свойствами целого организма и физиологически ему соответствует.
Тело простейших имеет микроскопические размеры и состоит из тех же компонентов, что и клетка многоклеточных — наружной мембраны, цитоплазмы, ядра и органоидов.
Наружная мембрана имеет типичное трехслойное строение, но за счет дополнительных образований у некоторых видов может достигать большой толщины и плотности. Цитоплазма подразделяется на два слоя: наружный и внутренний. Наружный слой (эктоплазма) более плотный, однородный и прозрачный, внутренний (эндоплазма) — зернистый, имеет более жидкую консистенцию. В эндоплазме находятся органоиды общего назначения — митохондрии, эндоплазматическая сеть, сетчатый аппарат и др.
Органоидами движения простейших служат: 1) псевдоподии или ложноножки, представляющие собой временные выросты цитоплазмы; 2) жгутики — постоянные органоиды, имеющие вид длинной тонкой нити, начинающейся обычно на переднем конце; 3) реснички — постоянные органоиды, представляющие собой многочисленные короткие нити.
Строение органоидов питания не одинаково и зависит от способа питания разных простейших. Большая часть простейших питается частицами твердой пищи. У таких организмов для переваривания пищи существует пищеварительная вакуоль — капля жидкости, содержащая пищеварительные ферменты, которая образуется при поступлении пищи в эндоплазму. Пищеварительная вакуоль окружает пищевую частицу и перемещается по телу простейшего.
Органоиды выделения представлены сократительной или пульсирующей вакуолью, имеющей вид небольшого пузырька, наполненного жидкостью, который периодически увеличивается, достигая определенного объема, а затем сокращается, выбрасывая жидкость наружу. Накопление и сокращение ритмически чередуются. Основная функция сократительной вакуоли — поддержание осмотического давления на постоянном уровне.
Большинство простейших имеет одно ядро, но существуют и многоядерные формы. Ядро имеет строение, характерное для эукариот. Морфология и формы деления ядер разнообразны, но, как
доказано в настоящее время, деление ядра в любом случае представляет собой митоз.
Размножение.Простейшие могут размножаться бесполым и половым путем. Бесполое размножение встречается как в виде деления на две части, так и в виде множественного деления. Половой процесс у большинства простейших представлен копуляцией, у инфузорий -конъюгацией.
Жизненный цикл. Чаще всего исходной стадией является зигота, затем следует бесполое размножение путем деления, затем образование половых особей и вновь образование зиготы.
Инцистирование. При неблагоприятных условиях внешней среды (повышение или понижение температуры, высыхание и т. д.) вегетативные формы простейших превращаются в цисты. Они перестают питаться, двигаться и покрываются толстой оболочкой; процессы обмена веществ при этом резко замедляются. При восстановлении благоприятных условий из цисты вновь выходит активная вегетативная форма. Инцистирование — защитная реакция, возникшая в процессе эволюции, обеспечивающая выживание в неблагоприятных условиях. Цисты могут сохраняться в течение длительного времени, месяцы и даже годы.
Классификация. Класс саркодовые (Sarcodina). 2. Класс жгутиковые (Flagellata s.Mastigophora). 3. Класс споровики (Sporozoa). 4. Класс инфузории (Infusoria).
Медицинское значение класса инфузорий.У человека паразитирует единственная инфузория – балантидий, которая обитает в пищеварительной системе. Имеются многочисленные виды инфузорий, паразитирующие в разных группах беспозвоночных животных.
3.22. КЛАСС САРКОДОВЫЕ. КЛАССИФИКАЦИЯ ПО ЛАТЫНИ. ОБЩАЯ ХАРАКТЕРИСТИКА. МЕДИЦИНСКОЕ ЗНАЧЕНИЕ.
Тип простейшие (Protozoa). Класс Саркодовые (Sarcodina).
Известно около 10000 видов саркодовых, 80% из них – обитатели моря.
Основная черта саркодовых – способность к образованию псевдоподий или ложноножек, которые служат для передвижения и захватывания пищи (представляют собой выросты цитоплазмы). В подразделении саркодовых на классы существенную роль играют органоиды движения.
Свыше 80% всех современных саркодовых являются обитателями моря. Некоторые виды живут в пресной воде, немногие приспособились к жизни в почве.Имеются также паразитические виды.
Класс саркодовых состоит из трех подклассов, весьма неравноценных по количеству относящихся к ним видов:
1. Корненожки (Rhizopoda) — около 2 тыс. видов;
2. Солнечники (Helizoa) —всего несколько десятков видов;
3. Лучевики (Radiolaria) — 7—8 тыс. видов.
Некоторые представители класса:
Пресноводная амёба (Amoeba proteus). Легко обнаруживается на поверхности ила из луж и прудов. Цитоплазма разделена на две части – эктоплазму и эндоплазму. Ложноножки – орган передвижения – образуются за счет перехода эктоплазмы из состояния геля в золь. Питается по мере передвижения J — т.е. когда она надвигается на водоросли или частицы органических веществ, она обтекает их и заглатывает. Переваривание происходит в пищеварительных вакуолях. (путём фагоцитоза). Реакция на внешнее раздражение – таксисы. Одно ядро. Осморегуляция – посредством выделения продуктов диссимиляции пульсирующими вакуолями. Размножаются бесполым путем – делением. При неблагоприятных условиях инцистируются.
Некоторые представители имеют скелет в виде раковины. (древнейшие – из их скелета образовался мел и известняки).
Паразитические саркодовые:
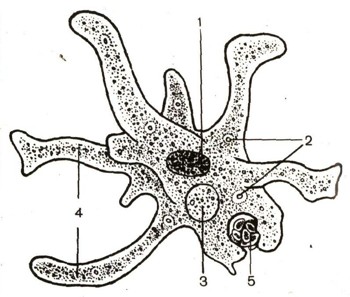 ДИЗЕНТЕРИЙНАЯ АМЕБА (Entamoeba histolytica). – возбудитель амебиаза или амебной дизентерии. Обитают в КИШЕЧНИКЕ ЧЕЛОВЕКА. Три формы, встречающихся в кишечнике у человека : 1. Крупная вегетативная, тканевая, патогенная (forma magna), 2. Мелкая, вегетативная форма, которая встречается чаще в просвете кишок (Forma minuta), 3. Цисты. Характерная черта цист дизентерийной амебы – 4 ядра в цисте. В организм человека дизентерийная амеба попадает в форме цист и в кишечнике она растворяется и из неё выходят 4 амебы мелкой формы. Эта форма питается бактериями. Это основная форма существования дизентерийной амебы. Если условия, необходимые для перехода в крупную форму, амебы инцистируются и выводятся во внешнюю среду, если подходят, то амеба увеличивается в размерах и приобретает способность выделять ферментативные белки,растворяющие тканевые белки. => они проникают в ткань, образуя кровоточащие язвы. Иногда заглатывают эритроциты. Попадая таким образом в кровь она может разноситься по всем остальным органам – печень и др. органы, вызывая абсцессы и язвы. Если условия, необходимые для существования формы магнум отсутствуют (период затухания болезни), то она превращается в форму минута. Условия, необходимые для перехода из малой формы в большую – переохлаждение, перегревание, недоедание, переутомление.
ДИЗЕНТЕРИЙНАЯ АМЕБА (Entamoeba histolytica). – возбудитель амебиаза или амебной дизентерии. Обитают в КИШЕЧНИКЕ ЧЕЛОВЕКА. Три формы, встречающихся в кишечнике у человека : 1. Крупная вегетативная, тканевая, патогенная (forma magna), 2. Мелкая, вегетативная форма, которая встречается чаще в просвете кишок (Forma minuta), 3. Цисты. Характерная черта цист дизентерийной амебы – 4 ядра в цисте. В организм человека дизентерийная амеба попадает в форме цист и в кишечнике она растворяется и из неё выходят 4 амебы мелкой формы. Эта форма питается бактериями. Это основная форма существования дизентерийной амебы. Если условия, необходимые для перехода в крупную форму, амебы инцистируются и выводятся во внешнюю среду, если подходят, то амеба увеличивается в размерах и приобретает способность выделять ферментативные белки,растворяющие тканевые белки. => они проникают в ткань, образуя кровоточащие язвы. Иногда заглатывают эритроциты. Попадая таким образом в кровь она может разноситься по всем остальным органам – печень и др. органы, вызывая абсцессы и язвы. Если условия, необходимые для существования формы магнум отсутствуют (период затухания болезни), то она превращается в форму минута. Условия, необходимые для перехода из малой формы в большую – переохлаждение, перегревание, недоедание, переутомление.
 Диагностика и пат.значение: выявляются нахождением в фекалиях человека цист с 4 ядрами. Бессимптомное протекание болезни возможно. Чаще – кровавый жидкий стул.
Диагностика и пат.значение: выявляются нахождением в фекалиях человека цист с 4 ядрами. Бессимптомное протекание болезни возможно. Чаще – кровавый жидкий стул.
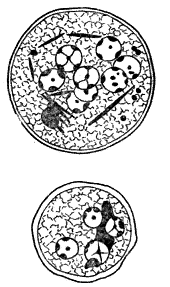
КИШЕЧНАЯ АМЕБА (Entamoeba coli). Непатогенна. Морфологически сходна с дизинтерийной. Так же образует вегетативные формы и цисты, но способности выделять ферментативные белки не имеет. Эритроциты не заглатывает. Циста содержит 8 ЯДЕР. Цисты кишечной амебы (коли) от дизентеррийной (хистолитика) отличаются размерами и числом ядер.
РИСУНОК. Цисты кишечной амебы (вверху) и дизентерийной амебы (внизу). В цистах видны ядра (у кишечной амебы их восемь, у дизентерийной — четыре) и хроматоидные тельца (по Кофоиду)
РОТОВАЯ АМЕБА (Entamoeba gingivalis). Первая паразитическая амеба, найденная у человека. Встречается в кариозных зубах и в белом налете, покрывающем зубы. Питается бактериями и лейкоцитами. Патогенное действие неясно.
3.23. КЛАСС ЖГУТИКОВЫЕ. КЛАССИФИКАЦИЯ ПО ЛАТЫНИ. ОБЩАЯ ХАРАКТЕРИСТИКА. ТРИПАНОСОМЫ И ВИДЫ ТРИПАНОСОМОЗОВ.
Известно около 6000-8000 видов. Жгутиковые (Flagellata ) делятся на растительные жгутиковые (Phytomastigina) или фитожгутиковые и животные жгутиковые (Zoomastigina) или зоожгутиковые. Представители первого подкласса имеют хроматофоры и питаются автотрофно, но в разных отрядах этого подкласса можно наблюдать постепенный переход от автотрофного питания к гетеротрофному. Представители второго подкласса не имеют хроматофоров и являются гетеротрофами. Многие из них (главным образом из отряда протомонадовых) ведут паразитический образ жизни. Ядро одно или несколько; покрыты оболочкой – пелликулой – определяющей форму клетки.
Отличительной особенностью класса является наличие одного или двух жгутиков, находящихся обычно на переднем конце животного. У основания жгутика имеется особый органоид — кинетопласт. Этот орган вырабатывает энергию для движения жгутика. Один из жгутиков, бывает, проходит вдоль тела и соединён с пелликулой посредством мембраны (ундулирующая мембрана), которая обеспечивает поступательно движение жгутиконосца. Размножение обычно бесполое, путём продольного деления. Встречается и половой процесс – копуляция – две особи сливаются, образуя зиготу.
К жгутиковым относятся лейшмании, трипаносомы, лямблия, трихомонады и хиломастикс.
ТРИПАНОСОМА (Trypanosoma gambiense) – возбудитель трипаносомоза – африканской сонной болезни.
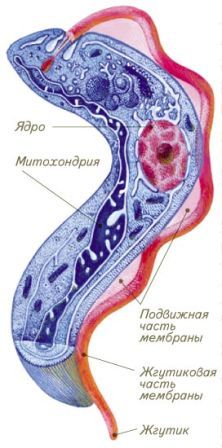
Обитает в плазме крови, в лимфе, лимфатических узлах, спинномозговой жидкости, тканях спинного и головного мозга. ВСТРЕЧАЮТСЯ в экваториальной Африке.
Тело изогнутое, сплющенное в одной плоскости, суженное на концах, один жгутик и ундулирующая мембрана. Питание осмотически пр-ит. РАЗМНОЖЕНИЕ происходит бесполым путем, продольным делением. Выделяют американский и африканский трипаносомоз (клоп, муха це-це).
Трипаносомы проходят две стадии развития — стадию трипомастиготы и стадию эпимастиготы. У возбудителя американского трипаносомозаописаны еще две стадии — стадия промастиготы и стадия амастиготы. Трипомастиготы паразитируют только в организме человека и только в организме насекомого. Поступая с кровью в организм мухи цеце (переносчик африканского трипаносомоза), трипаносомы попадают в слюнные протоки и железы, где превращаются в эпимастиготы. Через 2-5 дней эпимастиготы трансформируются в трипомастиготы, и мухи становятся заразными.
Попадая с кровью в организм триатомовых клопов (переносчики американского трипаносомоза), трипаносомы в желудке насекомого превращаются в эпимастиготы и размножаются здесь в течение нескольких дней. Затем они проходят в заднюю и прямую кишку, где возвращаются к трипомастиготной форме. С этого момента клопы становятся заразными.
ПО УЧЕБНИКУ ПРО АФРИКАНСКИЙ: ПЕРВАЯ ЧАСТЬ ЖИЗНЕННОГО ЦИКЛА – пищеварительный тракт мухи це-це (Glossina palpalis). В ней возбудитель трипаносомоза претерпевает ряд изменений, размножается, попадает он в муху при всасывании ею крови у зараженного человека. Цикл развития в составляет приблизительно 20 дней. Мухи, в теле которых содержится возбудитель трипаносомы в инвазионной стадиии, при укусе могут заразить человека.
У БОЛЬНОГО : мышечная слабость, истощение, умственная депрессия, сонливость. Болезнь может длиться 7-10 лет и может закончиться смертью. ДИАГНОСТИКА: исследование крови, лимфы, спинномозговой жидкости. ПРОФИЛАКТИКА: прием лекарств от заболевания, уничтожение разносчиков болезней (в д.с. мух це-це).
ИЗ ИНТЕРНЕТА:
Африканский трипаносомоз
Африканский трипаносомоз, или сонная болезнь, — трансмиссивная инфекция, передающаяся через кровососущих переносчиков, в данном случае — муху цеце, характеризующаяся периодами неправильной лихорадки, наличием кожной сыпи, местных отеков и лимфаденитов, развитием кахексии (истощения) и летаргии. Существует в двух формах — гамбийской и родезийской (восточно-африканской), отличающихся по эпидемиологии и особенностям клинического течения.
Американский трипаносомоз
Американский трипаносомоз — болезнь Чагаса. В 1907 г. Чагас обнаружил в кишечнике клопа трипаносому, в 1909 г. он выделил от больного трипаносому, идентичную найденной у клопа. Эти исследования положили начало изучению новой протозойной (вызываемой простейшими) инфекции.
Трипаносома имеет длину 15-20 мкм. В организме человека и млекопитающих животных трипаносомы проникают в клетки внутренних органов и тканей, где превращаются в амастиготы. Со временем амастиготы превращаются в S-образные трипомастиготы, которые поступают в кровь, но в ней не размножаются.
3.24. ЛЕЙШМАНИИ И ЛЕЙШМАНИОЗЫ. КЛАССИФИКАЦИЯ ПАРАЗИТОВ (ПО ЛАТЫНИ). ОСОБЕННОСТИ. ДИАГНОСТИКА И ПРОФИЛАКТИКА. ЗНАЧЕНИЕ РАБОТ П.А. ПЕТРИЩЕВОЙ, В.Л. ЯКИМОВА, П.Ф.БОРОВСКОГО.
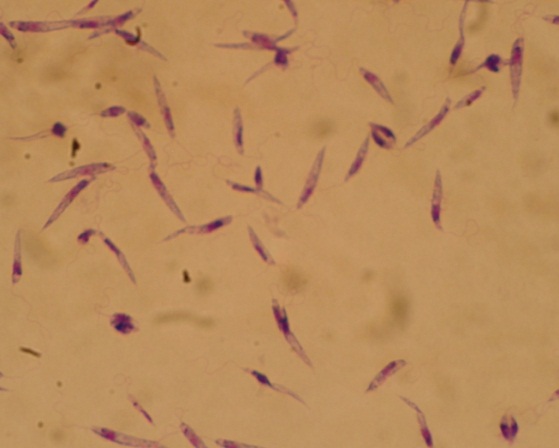
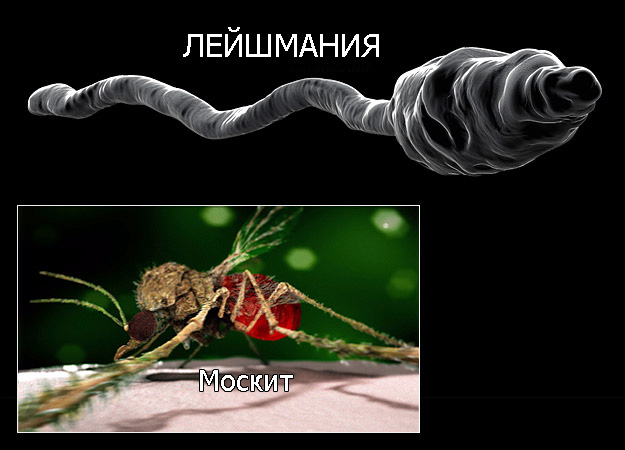 Лейшмании – возбудители лейшмаиоза – трансмиссивных заболеваний с природной очаговостью. Лейшмании открыты русским врачом Боровским П.Ф. в 1898 г. Все лешмании подразделяются на две группы видов: дерматотропные (локализующиеся в коже) и висцеротропные (локализующиеся во внутренних органах).
Лейшмании – возбудители лейшмаиоза – трансмиссивных заболеваний с природной очаговостью. Лейшмании открыты русским врачом Боровским П.Ф. в 1898 г. Все лешмании подразделяются на две группы видов: дерматотропные (локализующиеся в коже) и висцеротропные (локализующиеся во внутренних органах).
К ВОЗБУДИТЕЛЯМ КОЖНОГО ЛЕЙШМАНИОЗА относятся виды Leishmania tropica, Leishmania major и ряд других. ЛОКАЛИЗИРУЮТСЯ в клетках кожи.
Кожный лейшманиоз распространен в ряде стран Европы, Азии и Америки. В России лейшманиоз, называемся L. tropica ликвидирован, а L.major встречается в Закавказье и Средней Азии.
ОСОБЕННОСТИ МОРФОЛОГИЧЕСКИЕ. Лейшмании очень малы (2-4, иногда 8 мкм). Это внутриклеточное паразиты. В жизненном цикле проходят две стадии: безжгутиковая – овальной, округлой формы (в организме человека или других позвоночных), и жгутиковую, имеющую удлинённое тело (в организме насекомых-переносчиков). Жгутиковые формы образуются так же в искусственых культурах.
ПЕРЕНОСЧИКОМ ВОЗБУДИТЕЛЯ ЯВЛЯЮТСЯ МЕЛКИЕ НАСЕКОМЫЕ МОСКИТЫ (Phlebotomus). Естественным резервуаром L. tropica является человек, L. major – грызуны.
ПРИ КОЖНОМ ЛЕЙШМАНИОЗЕ образуются округлые, долго незаживающие изъязвления на открытых частях тела, преимущественно на лице. После заживления остаются зубцы. Иммунитет остается на них всю жизнь. Для лабораторной диагностики берут отделяемое из язвы и приготовляют мазки, которые микроскопируют.
ПРОФИЛАКТИКА заключается в личной защите от укусов москитов. Можно проводить прививки от лейшманиоза. А общественная профилактика – уничтожение москитов и грызунов.
ВИСЦЕРАЛЬНЫЙ ЛЕЙШМАНИОЗ. L. donovani, L. infantum.
Они сначала попадают в клетки кожи. При исследовании в световой микроскоп морфологически не отличаются от локализующихся в коже. Жизненный цикл такой же, как и у кожных, переносчики – москиты, заражающиеся от людей и собак, а в дикой природе от шакалов( и др. псовые) и грызунов.
Наиболее часто болеют дети до 12 лет. Повышенная температура, увеличенная селезёнка и печень, уменьшение содержания эритроцитов в крови.
Диагностика – пункция грудины, лимфатических узлов.
3.25. Класс Споровики. Классификация (по латыни). Общая характеристика отряда Кокцидий. Цикл развития. Диагностика и профилактика токсоплазмоза.
Тип Споровики (Sporozoa)
Отряд Грегарины (Gregarinida)
Отряд Кокцидии (Coccidia)
Отряд Кровяные споровики (Haemosporidia)
Тип Книдоспоридии (Cnidosporidia)
Класс Миксоспоридии (Myxosporidia)
Тип Микроспоридии (Microsporidia)
Тип Инфузории (Infusoria), или (Ciliata)
Известно около 1350 видов споровиков. Все споровики – паразиты. Общая черта – отсутствие у зрелых форм каких-либо органоидов движения, а так же сложный жизненный цикл со сменой полового и бесполого размножения в форме схизогонии. В одной из стадий развития споровиков образуются споры. Паразитический образ жизни сказался на упрощении их организации: они не имеют органов пищеварения и выделения. Питание, дыхание, выделение осуществляется всей поверхностью тела.
ОТРЯД КОКЦИДИЙ (Coccidia).
Паразиты эпителия кишечника и протоков печени кроликов, рогатого скота, многих домашних животных и диких птиц, насекомых и других животных. Особенно часто кокцидиоз поражает кроликов, вызывая их массовую гибель. Кокцидиозом болеют и люди.
Циклы развития кокцидии весьма сложны. Примером может служить цикл развития паразита кроликов эймерии (Elmeriastiedae).
Эймерия паразитирует в клетках эпителия протоков печени кроликов. Бесполое размножение этого споровика протекает по типу шизогонии. Паразит, находясь в клетке эпителия печени хозяина, растет, пока не достигнет зрелости, после чего его ядро рядом последовательных делений дает значительное число мелких ядер. Затем вокруг каждого из них обособляется участок цитоплазмы, что ведет к образованию новых мелких особей — мерозоитов.

Жизненный цикл кокцидий рода Eimeria характеризуется тремя периодами: 1) шизогонией, 2) гаметогонией и 3) спорогонией. Шизогония и гаметогония происходят в эпителиальных клетках кишечника и развиваются внутри клеток хозяина (эндогенно). Спорогония — во внешней среде, вне хозяина – экзогенно.
Так, кокцидий Eimeria media в стадии зрелой ооцисты попадают с водой или кормом в кишечник животного. Спорозоиты выходят из ооцисты и проникают в эпителиальные клетки кишечника, где растут и превращаются в шизонтов. Внутри такого шизонта образуются мелкие паразиты — мерозоиты. С развитием последних шизонт делится и разрушает эпителий стенки кишечника. Освободившиеся мерозоиты вновь проникают в эпителиальные клетки кишечника, образуя шизонты II, III, а у отдельных видов — IV и пятой генераций. Таким образом, шизогония этого рода КОКЦИДИЙ повторяется много раз. Затем бесполое множественное деление сменяется половым процессом — гаметогонией.
Сущность гаметогонии заключается в том, что последующие генерации шизонтов формируют мерозоиты, которые проникают в клетку хозяина и превращаются в одноядерные трофозоиты. В дальнейшем из одноядерных трофозоитов образуются макрогаметоциты и микрогаметоциты. Затем макрогаметоциты превращаются в макрогаметы. У микрогаметоцитов ядро делится, в результате образуются мелкие мужские клетки — микрогаметы. После образования макрогамет и микрогамет они копулируют (сливаются), образуя копулу или зиготу. Зигота окружается оболочкой и превращается в ооцисту. Ооцисты в зависимости от видов могут иметь самую разнообразную форму и величину, у всех у них двуконтурная оболочка и зернистая цитоплазма. Такие ооцисты выходят во внешнюю среду и проходят стадию спорогонии. Во внешней среде при наличии тепла, влаги и кислорода в ооцисте формируются четыре споры и в каждой из них по два спорозоита. С образованием в ооцисте спор и спорозоитов заканчивается спорогония. Такие ооцисты становится зрелыми, при попадании в организм восприимчивых животных и заражают их.