
Содержание
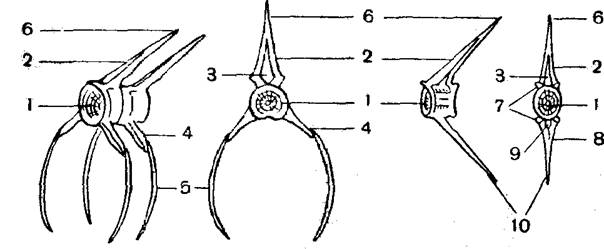
Б – то же костистой рыбы; В – поперечный разрез через жабру акулы; Г – то же костистой рыбы
1 – ротоглоточная полость; 2 – пищевод; 3 – челюстная дуга;
4 – подъязычная дуга; 5 – брызгальце; 6 – внутренние жаберные щели; 7 – наружные жаберные щели; 8 – жаберная крышка; 9 – жаберные дуги; 10 – жаберные лепестки; 11 – межжаберные перегородки.
На внутренней стороне жаберной крышки можно отметить остатки жабры, так называемую ложножабру, которая участия в газообмене не принимает.
Акт дыхания костистых рыб осуществляется за счет движения жаберной крышки: когда жаберная крышка приподнимается, тонкая кожистая жаберная перепонка, которая продолжается за край жаберной крышки, прижимается к жаберной щели. Это происходит под влиянием наружного давления воды. В результате в околожаберной полости образуется пространство с пониженным давлением. Это заставляет воду всасываться в околожаберную полость. При опускании крышки создается избыточное давление, и вода через наружные жаберные отверстия выталкивается наружу.
Кровеносная система костистых рыб (рис. 29), по сравнению с кровеносной системой хрящевых рыб, отличается некоторыми особенностями. Вместо артериального конуса появляется луковица аорты, которая является утолщением аорты и обладает, как и все сосуды, гладкой мускулатурой. Сохраняется по 4 пары приносящих и выносящих жаберных артерий (у хрящевых рыб их 5). Боковые вены исчезают. Только левая кардинальная вена распадается в левой почке на сеть капилляров, правая кардинальная вена идет через почку, не прерываясь (асимметрия воротной системы почек).
В головном отделе на брюшной стороне располагается сердце. Хорошо заметно темно-бордовое предсердие. Выше предсердия находится венозная пазуха, имеющая вид воронки и собирающая венозную кровь со всего тела. Снизу к предсердию примыкает ярко-красный желудочек. Разница в окраске предсердия и желудочка обусловлена толщиной стенок.
От желудочка отходит брюшная аорта, которая у основания расширяется в луковицу аорты.
Из брюшной аорты кровь по приносящим жаберным артериям направляется в жабры, где обогащается кислородом, затем, по выносящим жаберным артериям впадает в парные корни аорты и далее в спинную аорту. Сосуды, по которым артериальная кровь направляется к голове, называются сонными артериями.
По спинной аорте кровь идет по всему телу к хвостовой части, давая ответвления ко всем органам: желудку, кишечнику, половым органам, почкам, конечностям (плавникам). На теле вскрытой рыбы спинная аорта хорошо заметна между почками.
Венозная кровь возвращается в сердце по непарной хвостовой вене и по парным задним и передним кардинальным венам. Правая задняя кардинальная вена проходит через почки не разветвляясь, а левая дает сеть капилляров. Таким образом, воротная система почек развита только в левой почке. Задние кардинальные вены сливаются с передними кардинальными венами и образуют кювьеровы протоки.

Рисунок 29 – Схема кровеносной системы костистой рыбы
1 – предсердие; 2 – желудочек; 3 – луковица аорты; 4 – брюшная аорта; 5 – приносящие жаберные артерии; 6 – выносящие жаберные артерии;
7 – корни аорты; 8 – спинная аорта; 9 – сонные артерии; 10 – подключичные артерии; 11 – хвостовая вена; 12 – правая задняя кардинальная вена; 13 – левая задняя кардинальная вена; 14 – воротная система почек;
15 – подкишечная вена; 16 – воротная система печени; 17 – кювьеров проток; 18 – печеночная вена; 19 – передние кардинальные вены (в черный цвет окрашены сосуды с венозной кровью).
Подкишечная вена проходит через печень, распадаясь там на капилляры и образуя воротную систему печени. Из печени выходят печеночные вены, впадающие в венозную пазуху.
Органы выделения костистых рыб сходны с таковыми у хрящевых. Отличие заключается в том, что выделительная система костистых рыб не связана с органами размножения (рис. 30).
Длинные туловищные почки (мезонефрос) лежат по бокам позвоночника над плавательным пузырем. Мочеточниками служат вольфовы каналы, которые тянутся вдоль внутреннего края почек. У костистых рыб развит мочевой пузырь. Выделительная система имеет самостоятельное выделительное отверстие, где заканчиваются мочеточники.

Рисунок 30 – Мочеполовая система самца щуки
1 – почка; 2 – мочеточник; 3 – мочевой пузырь; 4 – наружное мочевое отверстие; 5 – семенник; 6 – семяпровод; 7 – наружное половое отверстие; 8 – кишечник; 9 – анальное отверстие.
Органы размножения представлены у самцов семенниками, у самок яичниками (рис. 30).
В отличие от хрящевых рыб, половые железы костистых рыб, как самцов, так и самок, имеют самостоятельные протоки – вторичные половые пути, которые не имеют ничего общего с вольфовыми и мюллеровыми каналами. По этим протокам половые продукты через половое отверстие выходят наружу. Таким образом, у самцов костистых рыб вольфов канал выполняет только одну функцию – выведения продуктов обмена, т.е. мочеточника. У самок мюллеров канал отсутствует.
В зависимости от сезона и возраста рыбы степень развития органов размножения может быть разная. У молодых экземпляров или особей, пойманных вне сезона размножения, половые органы развиты слабо. В период размножения размеры гонад сильно увеличиваются.
Головной мозг у костистых рыб имеет более примитивное строение, чем у хрящевых. Размеры его меньше, на крыше переднего мозга отсутствует нервное вещество, т.е. коры нет. Полости больших полушарий (боковые желудочки) не разделены перегородкой, передний мозг относительно мал. Наиболее развиты средний мозг и мозжечок.
Передний мозг представлен большими полушариями. Крыша их эпителиальная, не содержит мозгового вещества. Главную массу переднего мозга составляют полосатые тела, лежащие на дне.
Промежуточный мозг прикрыт большими полушариями. Если их раздвинуть в задней части, то можно обнаружить маленький булавовидный вырост – эпифиз. Это железа внутренней секреции.
Наибольшего развития достигает средний мозг, а именно, его зрительные доли.
Мозжечок также достаточно велик. Он налегает на продолговатый мозг.
Продолговатый мозг постепенно переходит в спинной. На продолговатом мозгу можно заметить ромбовидную ямку (четвертый мозговой желудочек).
От передней части больших полушарий отходят обонятельные доли, а от них – обонятельные нервы, которые разветвляются в носовых капсулах.
Скелет костистых рыб
Теоретические сведения:
8.1 Общая характеристика
8.2 Строение скелета
Общая характеристика
Скелет костистых рыб, в отличие от хрящевых, почти целиком костный. Он состоит из костей двух типов: хондральных, или замещающих костей, возникающих путем последовательной замены хряща костной тканью, и покровных, или кожных костей, образующихся в соединительнотканном слое кожи независимо от хрящевого скелета.
Покровные кости обычно располагаются поверхностно и имеют вид пластинок. Покровные кости не имеют хрящевых предшественников, и их образование приводит к появлению новых элементов скелета, а следовательно, к его усложнению.
Осевой скелет:
Представлен хорошо развитыми костными амфицельными позвонками. В телах позвонков и между ними проходит четковидная хорда. Позвоночный столб представлен туловищным и хвостовым отделами, строение которых аналогично хрящевым рыбам. Позвонки соединены с помощью суставных отростков, расположенных у основания верхних дуг.
Череп:
а) Мозговой череп – характерно наличие большого количества основных и покровных костей.
— в затылочном отделе 4 затылочные кости: основная затылочная, 2 боковые и верхняя затылочная кости.
— боковой отдел образован 5 ушными костями, 3 глазничными костями (глазоклиновидная, основная и боковая клиновидная), 2 обонятельными костями (непарная средняя обонятельная и боковые парные обонятельные). Все эти кости основные: развиваются путем окостенения хряща.
— крыша мозгового черепа образована покровными костями: парными носовыми, лобными и теменными костями.
— дно мозгового черепа образовано 2 непарными кожными костями: парасфеноидом и сошником с зубами.
б) Висцеральный череп – образован челюстной, подъязычной, 5 парами жаберных дуг и скелетом жаберной крышки.
— челюстная дуга делится на первичные челюсти – окостенение хрящевых элементов челюстной дуги, и вторичные челюсти – покровные кости, укрепляющие челюсти. Из небно-квадратного хряща (верхняя челюсть) образуются 3 основные кости: небные (с зубами), задние крыловидные и квадратная. Между ними располагаются покровные наружные и внутренние крыловидные кости. Из меккелева хряща (нижняя челюсть) образуется замещающая сочленовная кость, образующая с квадратной костью челюстной сустав. Вторичные челюсти представлены в верхней челюсти предчелюстными и верхнечелюстными костями с зубами; в нижней челюсти – зубной и угловой костями.
— подъязычная дуга образована основными костями: гиомандибуляре, гиоидом и непарной копулой. Для костных рыб характерна гиостилия.
— скелет жаберной крышки представлен 4 покровными костями: предкрышечной, крышечной, межкрышечной и подкрышечной.
— жаберных дуг 5 пар. Первые 4 образованы 4 парными элементами, соединенными снизу копулами (они несут жабры). Последняя жаберная дуга не несет жабр и состоит из 2 парных элементов, к которым могут быть причленены глоточные зубы (у некоторых).
Скелет парных конечностей и их поясов:
Парные конечности представлены грудными и брюшными плавниками. Различают 2 типа парных плавников:
а) бисериальный тип – плавники имеют центральную расчлененную ось, к которой попарно прикрепляются членики радиалий (лопастеперые и двоякодышащие);
б) унисериальный тип – радиалии прикрепляются только с одной стороны центральной оси (кистеперые рыбы).
У лучеперых рыб базальные элементы плавников редуцируются, радиалии прикрепляются непосредственно к поясу, к радиалиям прикрепляются лепидотрихии (кожные костные лучи, поддерживающие лопасть плавника).
Плечевой пояс состоит из первичных и вторичных элементов. Первичный пояс представлен окостеневшими лопатками и коракоидом. Вторичный пояс представлен крупным клейтрумом, который посредством супраклейтрума присоединяется к затылочному отделу черепа.
Скелет собственно грудных плавников представлен одним рядом радиалий, к которому крепятся лепидотрихии.
Тазовый пояс представлен хрящевой или костной пластинкой, лежащей в толще мускулатуры, к которой через ряд радиалий крепятся лепидотрихии брюшных плавников.
Скелет непарных конечностей:
Спинные плавники образованы лепидотрихиями, скелетной основой которых являются птеригофоры, погруженные в мускулатуру и нижними концами соединенные с остистыми отростками позвонков.
Хвостовой плавник бывает:
а) протоцеркальный – симметричное строение, хорда проходит по середине плавника (личинки рыб).
б) гетероцеркальный – аналогичен хрящевым рыбам (у осетрообразных).
в) гомоцеркальный – равнолопастной, верхняя и нижняя лопасти одинаковы, но осевой скелет заходит в верхнюю лопасть (большинство костистых рыб).
г) дифицеркальный – однолопастной. Осевой скелет проходит по середине плавника (двоякодышащие и кистеперые рыбы).
Скелетной основой хвостового плавника являются расширенные отростки концевых позвонков – гипуралии, лопасть плавника поддерживают лепидотрихии.
Строение скелета
Череп костистых рыб (рис. 31) характеризуется сложностью строения, возникшей в результате появления большого количества замещающих и покровных костей.
Затылочный отдел представлен четырьмя затылочными костями, которые окружают затылочное отверстие. Сверху лежит верхняя затылочная кость, по бокам – боковые затылочные кости, снизу расположена основная (нижняя) затылочная кость. Все кости этого отдела хондрального происхождения, так как замещают затылочный отдел хрящевых рыб.

Рисунок 31 – Схема строения черепа костистой рыбы
1 – нижняя затылочная кость; 2 – боковая затылочная кость;
3 – верхняя затылочная кость; 4 – ушные кости; 5 – основная клиновидная кость; 6 – крылоклиновидная кость; 7 – глазоклиновидная кость; 8 – средняя обонятельная кость; 9 – боковая обонятельная кость;
10 – теменная кость; 11 – лобная кость; 12 – носовая кость;
13 – парасфеноид; 14 – сошник; 15 – небная кость; 16 – квадратная кость; 17 – крыловидные кости; 18 – межчелюстная кость;
19 – верхнечелюстная кость; 20 – сочленовная кость; 21 – зубная кость; 22 – угловая кость; 23 – гиомандибуляре; 24 – симплектикум;
25–29 – жаберные дуги; 30 – гиоид; 31 –копула (точками помечены замещающие кости).
Бока черепа представлены пятью ушными костями, расположенными в задней части черепа, которые заместили слуховую капсулу хрящевых рыб. Заднюю часть глазницы выстилает крылоклиновидная кость. Глазоклиновидная кость имеется только у низших костистых рыб. Клиновидные кости хондрального происхождения, так как развиваются на месте глазницы хрящевых рыб. В передней части глазницы расположена слезная кость, пронизанная слезным каналом. Глазничное кольцо, ограничивающее глазную впадину, образовано предглазничными костями. Слезная кость и предглазничные кости покровного происхождения.
Крыша черепа образована рядом костей. Парные теменные кости лежат по бокам верхней затылочной кости. Крупные лобные кости соединяют затылочный отдел со средней обонятельной костью, находящейся в передней части черепа. По бокам от нее находятся парные носовые и лежащие позади них боковые обонятельные кости. Кости крыши черепа покровного происхождения за исключением средней и боковых обонятельных костей, которые имеют смешанное происхождение.
Дно черепа подстилает палочковидная кость – парасфеноид. Впереди он заканчивается сошником, несущим зубы. Обе кости покровного происхождения.
Висцеральный череп представлен рядом дуг (как и у хрящевых рыб) (рис. 31).
Челюстная дуга костистых рыб сильно отличается от челюстной дуги хрящевых. На месте небно-квадратного хряща (первичной верхней челюсти) хрящевых рыб возникают небная и квадратная кости. Небная кость направлена вперед и снабжена зубами. Квадратная кость смещена назад и функцию челюсти не несет.
Между небной и квадратной костями появляются три крыловидных кости. Две из них (покровного происхождения) разделяют небную и квадратную кости. Крупная, округлая по форме задняя крыловидная кость (смешанного происхождения) лежит позади первых двух. Она образовалась в результате окостенения части небно-квадратного хряща.
Кроме того, появляется вторичная верхняя челюсть, образованная парными предчелюстной (межчелюстной) и верхнечелюстной костями. Эти кости обычно несут зубы и служат для захвата и удержания пищи, то есть выполняют роль настоящих челюстей.
Нижняя челюсть костистых рыб представлена покровными зубной и угловой костями. В заднем углу нижней челюсти расположена хондральная сочленовная кость, заместившая меккелев хрящ.
Позади челюстной дуги находится подъязычная дуга, схожая с таковой хрящевых рыб. Ее верхний элемент – гиомандибуляре соединяет мозговой череп с челюстями и служит подвеском. Таким образом, у костистых рыб, так же как и у хрящевых, имеет место гиостилия. Ниже гиомандибуляре лежит небольшая косточка – симплектикум и второй элемент подъязычной дуги – гиоид. Обе половины подъязычной дуги соединяет копула.
У костистых рыб четыре пары жаберных дуг. Каждая дуга состоит из четырех элементов. Пятая жаберная дуга значительно редуцирована и представлена лишь одним элементом. У растительноядных рыб, например карповых, на ней сидят глоточные зубы.
Все кости подъязычной дуги и жаберных дуг хондрального происхождения.
Жаберные дуги сверху прикрывает жаберная крышка, состоящая из четырех покровных костей. Книзу от тонкой плоской крышечной кости лежит подкрышечная кость. Впереди крышечной кости находится зазубренная серповидная предкрышечная кость, а несколько ниже – межкрышечная кость.
Позвоночный столб у костистых рыб, так же как у хрящевых, состоит из амфицельных (двояковогнутых) позвонков. Между ними залегает хорда, проходящая сквозь тела позвонков в виде тяжа и в целом имеющая четковидную форму.
А Б
Рисунок 32 – Строение туловищного (А) и хвостового (Б) позвонков костистой рыбы (сбоку и спереди)
1 – тело позвонка; 2 – верхняя дуга; 3 – спинномозговой канал;
4 – поперечные отростки; 5 – ребра; 6 – верхний остистый отросток; 7 – сочленовные отростки; 8 – нижняя дуга; 9 – гемальный канал;
10 – нижний остистый отросток.
Туловищный позвонок (рис. 32) состоит из тела позвонка, верхней и нижней дуги. Верхняя дуга ограничивает спинномозговой канал и заканчивается верхним остистым отростком. Нижняя дуга разрастается в стороны в виде двух поперечных отростков, к которым причленяются ребра.
У хвостовых позвонков хорошо выражены нижние дуги, которые образуют гемальный канал и заканчиваются нижним остистым отростком. По гемальному каналу проходят кровеносные сосуды.
У костистых рыб на позвонках появляются сочленовные поверхности для более подвижного соединения позвонков. К хорошо развитым верхним и нижним остистым отросткам прикрепляется мускулатура.
Позвоночный столб у костистых рыб, так же как и у хрящевых, неподвижно соединен с черепом.
Пояса конечностей представлены плечевым и тазовым поясами.

Рисунок 33 – Плечевой пояс костистой рыбы с грудным плавником
1 – лопатка; 2 – коракоид; 3 – клейтрум; 4 – надклейтрум;
5 – заднетеменная кость; 6 – радиалии; 7 – лучи плавника (лепидотрихии).
Плечевой пояс (рис. 33) представлен костями хондрального происхождения – лопаткой и коракоидом (врановой костью). Лопатка уплощена, с отверстием посередине, коракоид серповидно изогнут. Они образуют первичный плечевой пояс.
Сверху к этим костям прикрепляется кость покровного происхождения клейтрум – большая серповидная кость с вытянутым, заостренным отростком. Через две маленькие покровные косточки – надклейтрум и заднетеменную кость – плечевой пояс неподвижно крепится к черепу. Это кости вторичного пояса.
Тазовый пояс прост и имеет форму треугольной кости (рис. 34).

Рисунок 34 – Тазовый пояс костистой рыбы с брюшным плавником
1 – тазовая кость; 2 – лучи плавника (лепидотрихии).
Парные плавники, как и у хрящевых рыб, представлены грудными и брюшными плавниками. Грудной плавник (рис. 33) заметно упрощен. Базалии отсутствуют, имеются лишь радиалии, от которых отходят кожные плавниковые лучи (лепидотрихии).
Брюшной плавник (рис. 34) не имеет ни базалий, ни радиалий. Лучи плавника (лепидотрихии) прикрепляются непосредственно к тазу.
Непарные плавники представлены спинными, хвостовым и анальным плавниками.
Непарный спинной плавник состоит из кожных плавниковых лучей. Они могут быть твердыми или мягкими. Основания их представлены птеригофорами, имеющими связь с верхними остистыми отростками туловищных позвонков. Такое же строение имеет анальный (подхвостовой) плавник.
Хвостовой плавник у костистых рыб гомоцеркального типа (рис. 35), т.е. равнолопастной. Однако позвоночник заходит в верхнюю лопасть. Такой плавник произошел из гетероцеркального плавника путем разрастания брюшной лопасти.
Тема 8. СКЕЛЕТ КОСТИСТОЙ РЫБЫ
Материал и оборудование. Смонтированный на картонных планшетах скелет костистой рыбы. Позвонки туловищного и хвостового отделов; грудные и брюшные плавники с поясами; вареные головы костистых рыб (по одному набору на 2-3 студентов). Таблицы: Скелет костистой рыбы; Строение туловищного и хвостового позвонков; Мозговой и висцеральный скелет головы; Скелет конечностей и их поясов; Строение непарных плавников. Ванночка, марлевые салфетки, пинцет, препаровальные иглы, лист ватмана, клей (по одному набору на 2-3 студентов).
Вводные замечания. Скелет костистых рыб состоит из костей хондральных, или первичных (образовались путем окостенения хряща), и покровных, или вторичных (образовавшихся в коже). В отличие от хрящевых ганоидов скелет целиком костный, жаберная крышка состоит из четырех костей; рострум отсутствует.
Осевой скелет представлен не хрящевой полой трубкой с хордой внутри, а амфицельными позвонками с верхними и нижними дугами. В туловищном отделе к последним прикрепляются ребра, прикрывающие полость тела не только сверху, но и с боков.
Задание. При выполнении работы нужно рассмотреть мозговой череп: крышу и дно черепа; обонятельный отдел; глазничный отдел; слуховой отдел; затылочный отдел; висцеральный череп; челюстную дугу: верхнюю и нижнюю челюсти; подъязычную дугу; жаберные дуги; жаберную крышку.
В осевом скелете нужно рассмотреть туловищный и хвостовой отделы позвоночника; туловищный позвонок и его строение; хвостовой позвонок и его строение; пояса конечностей: плечевой пояс и тазовый пояс; парные плавники; непарные плавники.
Изучение строения скелета рекомендуется начинать с самой сложной структуры – черепа. По рисункам, таблицам и готовым препаратам рассматривается общее расположение костей черепа. Следующий этап работы – разбор висцерального и мозгового черепа. Каждая кость извлекается из черепа, освобождается от тканей, промывается, высушивается и раскладывается в определенном порядке на листе ватмана. Поскольку плечевой пояс топографически связан со скелетом головы, допустимо рассматривать его вместе со скелетом черепа и также на приотваренном препарате. Практические рекомендации по разбору черепа даны в приложении к этой теме.
Расположенные по отделам кости висцерального и мозгового черепа, плечевого пояса (после проверки выполненной работы преподавателем) подключают на лист и подписывают.
Закончив работу со скелетом головы, следует перейти к изучению осевого скелета и скелета тазового пояса и плавников по макетам.
Строение скелета рассматривается на примере судака (окуня). Для сравнения приводятся некоторые особенности в строении скелета щуки, карпа, налима.
Скелет черепа. У костистых, так же как и у других рыб, череп делится на мозговой череп (neurocranium) и висцеральный (splanchnocranium).
Мозговой череп судака (окуня) тропибазального типа: с узким основанием и сближенными стенками глазниц (рис. 42). В нем можно выделить крышу черепа, дно, обонятельный, глазничный, слуховой и затылочный отделы. Крыша и дно черепной коробки образованы покровными костями. Крышу составляют 3 пары костей: носовые (nasale); лобные (frontale); теменные (parietale). Дно черепа образовано двумя непарными костями: лежащим впереди сошником (vomer), несущим зубы, и большим парасфеноидом (parasphenoideum).
В области обонятельного отдела имеется непарная средняя обонятельная кость (mesethmoideum) и парные боковые обонятельные кости (ectoethmoideum). В обонятельном отделе сохраняется много хряща, что делает хорошо заметными границы между обонятельными костями.
 Рисунок 42 – Осевой череп судака:
Рисунок 42 – Осевой череп судака:
а – вид сбоку; б – вид сверху; в – вид сзади; 1 – средняя обонятельная кость; 2 – боковая обонятельная кость; 3 – носовая кость; 4 – лобная кость; 5 – боковая клиновидная кость; 6 – клиновидноушная кость; 7 – крыловидноушная кость; 8 – теменная кость; 9 – верхнеушная кость; 10 – верхнезатылочная кость; 11 – боковая затылочная кость; 12 – основная затылочная кость; 13 – заднеушная кость; 14 – переднеушная кость; 15 – парасфеноид; 16 – основная клиновидная кость; 17 – сошник.
Глазничный отдел осевого черепа образован клиновидными костями, или сфеноидами. Среди них: парные боковые клиновидные (laterosphenoideum) и основная клиновидная (basisphenoideum). Глазница окружена кольцом тонких окологлазничных косточек (orbitale), передняя из которых наибольшая и называется слезной (lacrimale), лежащие снизу и сзади в количестве 4-5 – подглазничными (suborbitale). Слуховой отдел представлен пятью ушными костями с каждой стороны. Переднеушная кость (prooticum) большая, занимает переднюю часть ушного отдела. В области этой кости находится лабиринт и отолиты (их можно обнаружить в месте соприкосновения переднеушной кости с основной затылочной костью). В ней же есть отверстие для выхода тройничного нерва. 3аднеушная кость (opisthoticum) лежит позади переднеушной. Кроме этих костей в состав слухового отдела входят: клиновидноушная (sphenoticum), крыловидноушная (pteroticum) и верхнеушная (epioticum).
Затылочный отдел состоит из четырех костей, окаймляющих большое затылочное отверстие основной (basioccipitale), с которой сочленяется позвоночник; двух боковых (exocciptitaleoccipitale laterale) и верхней (supraoccipitale). Последняя несет гребень для прикрепления мышц.
Висцеральный скелет представлен 7 висцеральными дугами: челюстной, подъязычной и пятью парами жаберных дуг, из которых последняя сильно редуцирована. Система дуг представляет собой большое число отдельных окостенений, действующих комплексно по типу сложных рычагов. Весь этот комплекс соединенных друг с другом костей прикреплен к черепной коробке с помощью подвеска (гиомандибуляре). Таким образом, у костистых, так же как у хрящевых ганоидов, имеет место гиостилия.
Челюстная дуга состоит из верхней и нижней челюстей. Первичная верхняя челюсть, гомологичная небно-квадратному хрящу пластинчато-жаберных, образована парными несущими зубы небными (palatinum) костями, тремя крыловидными – наружной (ectopterygoideum), внутренней (entopterygoideum), задней (metapterygoideum) – и квадратной (quadratum) (рис. 43). Небная кость имеет смешанное происхождение; внутренняя и наружная крыловидные развиваются как покровные на небно-квадратном хряще, задняя крыловидная и квадратная – хондральные кости.
 Рисунок 43 – Висцеральный отдел черепа судака:
Рисунок 43 – Висцеральный отдел черепа судака:
1 – небная кость; 2 – наружная крыловидная кость; 3 – внутренняя крыловидная кость; 4 – задняя крыловидная кость; 5 – квадратная кость; 6 – предчелюстная кость; 7 – верхнечелюстная кость; 8 – сочленовая кость; 9 – зубная кость; 10 – угловая кость; 11 – гиомандибуляре; 12 – симплектикум; 13 – палочковидная кость; 14 – верхнеподъязычная кость; 15 – среднеподъязычная кость; 16 – нижнеподъязычная кость; 17 – язычная кость; 18 – заднеподъязычная кость; 19 – лучи жаберной перепонки.
Вторичная верхняя челюсть – хватательный аппарат, который состоит из покровных костей парных предчелюстных, или межчелюстных (praemaxillare), и верхнечелюстных (maxileare). Предчелюстная кость судака (окуня) несет зубы. Верхнечелюстная кость лежит над предчелюстной; ее задний конец расширен в виде тонкой пластинки.
Нижняя челюсть образована тремя парными костями: хондральной сочленовной (articulare), соединяющейся с квадратной костью и гомологичной меккелеву хрящу пластиножаберных, и двумя покровными – зубной (dentale) и угловой (аngulare), прикрепляющейся к заднему углу сочленовной. Зубная кость несет зубы.
Подъязычная дуга (arcus hyoideus) состоит из первичных костей. Верхний элемент ее – подъязычно-челюстная кость (hyomandibulare) широким верхним концом причленяется к слуховому отделу осевого черепа. Маленькая (добавочная) косточка (symplecticum) отходит от нижнего края гирмандибуляре и соединяет его с квадратной костью (гиостилия), а палочковидная (interhyale-stylohyale) связывает гиомандибуляре с гиоидами.
Нижний отдел подъязычной дуги представлен сложным гиоидом (hyoideum), состоящим из четырех частей: верхнеподъязычной (epihyale), среднеподъязычной (ceratohyale) и двух маленьких нижнеподъязычных (hypohyale). Нижние элементы гиоидной дуги на вентральной стороне соединены непарной подъязычной, или язычной (basihyale, glossohyale), костью, выполняющей функцию языка. От вентральной стороны гиоида отходит непарная заднеподъязычная, или горловая кость (urohyale). От верхней и среднеподъязычной частей гиоида отходят лучи жаберной перепонки (radii branchiostegi). Число их является систематическим признаком нeкоторых костистых рыб.
Жаберных дуг (arcus branchialis) у судака (окуня) пять пар, но последняя, пятая, сильно редуцирована. Первые 3 пары состоят из четырех косточек: глоточно-жаберные (pharyngobranchiale), верхнежаберные (epibranchiale), среднежаберные (ceratobranchiale) и нижнежаберные (hypobranchiale) (рис. 44). Все они подвижно сочленены между собой, а снизу примыкают к непарной основной жаберной косточке (basibranchiale), или копуле (copula). В четвертой паре жаберных дуг отсутствует нижнежаберная косточка, в 5-й сохраняются только среднежаберная косточка и непарная копула. На 5-й дуге у судака (окуня) находятся мелкие нижнеглоточные зубы. На верхних глоточно-жаберных косточках расположены мелкие верхнеглоточные зубы.
 Рисунок 44 – Жаберный аппарат судака:
Рисунок 44 – Жаберный аппарат судака:
1 – глоточно-жаберные кости; 2 – верхнежаберные кости; 3 – среднежаберные кости; 4 – нижнежаберные кости; 5 – копула.
Жаберная крышка образована четырьмя покровными костями: крышечной (operculum), подкрышечной (suboperculum), межкрышечной (interoperculum) и предкрышечной (praeoperculum) с зазубренным краем.
 У щуки мозговой череп тропибазального типа, сохраняет большое количество хряща, особенно в обонятельном отделе (рис. 45).
У щуки мозговой череп тропибазального типа, сохраняет большое количество хряща, особенно в обонятельном отделе (рис. 45).
Рисунок 45 – Осевой череп щуки:
а – вид сверху; б – вид снизу; 1 – предобонятельная кость; 2 – боковая обонятельная кость; 3 – боковая клиновидная кость; 4 – клиновидноушная кость; 5 – крыловидноушная кость; 6 – переднеушная кость; 7 – верхнезатылочная кость; 8 – верхнеушная кость; 9 – боковая затылочная кость; 10 – основная затылочная кость.
Носовые кости сильно вытянуты и на большом протяжении плотно соединяются с вытянутыми отростками лобных костей. В обонятельном отделе отсутствует непарная средняя обонятельная кость, но по бокам передней части рыла выступают из-под покровных носовых костей небольшие предобонятельные кости (praeethmoideum). Боковые обонятельные кости располагаются по бокам основания рострума. Отсутствует основная клиновидная кость. В окологлазничном кольце имеются две пары надглазничных косточек. Одна – округлой формы, вторая – вытянутая, передним концом она соприкасается с носовой костью, а медиальным краем – с отростком лобной. Ее можно считать предлобной костью (praefrontale). В слуховом отделе нет заднеушной кости. Верхнезатылочная кость крупная, имеет округлую форму; ее края прикрыты теменными костями, так что кость выглядит как узкий отросток между ними.
В висцеральном скелете следует отметить форму небной кости: она вытянута в длину, уплощена, снабжена щеткой зубов на нижней поверхности. Предчелюстные кости короткие и не соприкасаются, а лежат по бокам от передней части рострума и снизу усажены мелкими зубами. Верхнечелюстная кость длинная, является основной костью вторичной верхней челюсти; к ее переднему концу прикрепляется предчелюстная. Хорошо развиты верхнеглоточные зубы, а 5-я редуцированная жаберная дуга несет мелкие нижнеглоточные зубы.
У карпа мозговой череп платибазального типа: с широким основанием, глазницы отделены одна от другой (рис. 46). Кости черепа плотно соединены между собой; количество хряща незначительное. Носовые кости короткие, округлые, расположены по бокам от
 Рисунок 46 – Осевой череп карпа:
Рисунок 46 – Осевой череп карпа:
а – вид сбоку; б – вид сверху; в – вид сзади; 1 – лобная кость; 2 – теменная кость; 3 – сошник; 4 – парасфеноид; 5 – средняя обонятельная кость; 6 – боковая обонятельная кость; 7 – глазоклиновидная кость; 8 – боковая клиновидная кость; 9 – переднеушная кость; 10 – заднеушная кость; 11 – клиновидноушная кость; 12 – крыловидноушная кость; 13 – верхнеушная кость; 14 – основная затылочная кость; 15 – боковая затылочная кость; 16 – верхнезатылочная кость; 17 – фонтанели.
сошника и средней обонятельной кости с широкими крыловидными образованиями. Сошник короткий и не несет зубов. В глазничном отделе имеются глазоклиновидные кости (orbitosphenoideum). Вместе с боковыми клиновидными они образуют межглазничную перегородку, характерную для платибазального типа черепа. На середине верхнего края глазницы лежит одна пара надглазничных косточек. Базисфеноид отсутствует. Среди ушных костей своими размерами выделяется переднеушная, отделяющаяся от боковой затылочной кости отверстием. Заднеушная кость, напротив, очень мала, располагается между боковой затылочной и крыловидноушной. Последняя составляет верхний край большой впадины в слуховой области основная затылочная кость снизу несет мощный отросток, на нем лежит жерновок. Основание отростка имеет отверстие, через которое проходит спинная аорта. Боковые затылочные кости – с большими латеральными крыльями. К их средней стороне, через клейтрум, причленяется пояс передних конечностей. Эти кости имеют два крупных: отверстия – фонтанели. Таким образом, в затылочном отделе карпа три отверстия, а не одно, как у других рыб. Фонтанели служат для прохождения затылочных нервов, а также, видимо, сообщения полости черепа с внечерепными лимфатическими пространствами.
В висцеральном скелете карпа можно отметить, ряд особенностей. Небная кость короткая, состоит из трех отростков. Предчелюстные кости срастаются своими верхними концами. Верхнечелюстные – широкие, сложной формы. Все кости верхней и нижней челюстей не несут зубов. Среднежаберные кости увеличены, и на них расположены трехрядные нижние глоточные зубы. Верхнеглоточные зубы отсутствуют.
У налима, как у судака и щуки, мозговой череп тропибазального типа. Хряща относительно мало, он играет подчиненную роль. Череп сбоку низкий; основание его прямое, и лишь задний конец основной затылочной кости несколько изогнут вниз (рис 47).
 Рисунок 47 – Череп налима:
Рисунок 47 – Череп налима:
а – вид сбоку; б – вид сверху; в – вид снизу; г – вид сзади; 1 – средняя обонятельная кость; 2 – боковая обонятельная кость; 3 – лобная кость; 4 – клиновидноушная кость; 5 – теменная кость; 6 – крыловидноушная кость; 7 – верхнеушная кость; 8 – верхнезатылочная кость; 9 – боковая затылочная кость; 10 – основная затылочная кость; 11 – парасфеноид; 12 – заднеушная кость; 13 – переднеушная кость; 14 – боковая клиновидная кость; 15 – сошник.
Головка сошника сильно развита, снизу черепа она имеет вид подковы и вся покрыта мелкими щетиновидными зубами. Рукоятка сравнительно узкая. Парасфеноид широкий, нижний край его на всем протяжении закругленный. Череп сверху довольно широкий у заднего конца, сужающийся к переднему. Череп сзади с почти невыдвигающимися над ним гребнями верхнезатылочной кости. Нет орбитосфеноида и базисфеноида. В центре заднеушной кости находится отверстие для прохождения нерва. Отолиты очень крупные.
Висцеральный скелет отличается следующим. Небные кости короткие и не несут зубов. На предчелюстных костях и костях нижней челюсти слабые щетинковидные зубы. Верхнечелюстная кость узкая. Хорошо развиты многочисленные верхнеглоточные и нижнеглоточные зубы. Крышечная кость узкая и удлиненная, нижний край ее с выемкой.
У лосося, щуки и других рыб некоторые из перечисленных костей могут заменяться покровными, и тогда они носят иные названия. В обонятельном отделе средняя обонятельная кость может развиваться как покровная. Тогда она называется верхнеобонятельной (supraethmoideum). Если боковые обонятельные кости являются покровными, то они называются предлобными (praefrontale). Из окостенений слуховой капсулы постоянно хрящевыми костями сохраняются переднеушная, верхнеушная и заднеушная. Клиновидноушная и крыловидноушная у некоторых рыб могут быть покровными. В таком случае клиновидноушная называется заднелобной (postfrontale), крыловидноушная – чешуйчатой (squamosus).
Осевой скелет (позвоночник). У судака (окуня) он образован костными амфицельными (двояковогнутыми) позвонками, между ними сохраняются остатки хорды. Общее количество позвонков у судака 42-48, у окуня – 39-42. В осевом скелете выделяют два отдела: туловищный и хвостовой. Позвонки туловищного и хвостового отделов имеют различное строение, это можно увидеть на рисунке 48. Каждый позвонок имеет тело позвонка (corpus vertebrae), от верхней (спинной) части которого отходят верхние невральные
 Рисунок 48 – Строение туловищного (а) и хвостового (б) позвонков костистой рыбы (вид спереди и сбоку):
Рисунок 48 – Строение туловищного (а) и хвостового (б) позвонков костистой рыбы (вид спереди и сбоку):
1 – тело позвонка; 2 – невральная дуга; 3 – верхний остистый отросток; 4 – сочленовый отросток; 5 – парапофиз; 6 – гемальная дуга; 7 – нижний остистый отросток.
дуги (arcus neuralis), замыкающиеся верхним остистым отростком (processus spinosus). Основания верхних дуг образуют сочленовные отростки – зигапофизы (sygapophysis). Снизу с боков от тела позвонка отходят поперечные отростки – парапофизы (parapophysis). В туловищных позвонках к ним причленяются ребра (costa). В позвонках хвостового отдела поперечные отростки срастаются, образуя нижнюю гемальную дугу (arcus haemalis), замыкающуюся нижним остистым отростком (processus spinosus inferior). В невральном канале располагается спинной мозг, в гемальном – хвостовая артерия и хвостовая вена. Начало гемального канала служит границей между туловищным и хвостовым отделами, а количество позвонков в них является важным систематическим признаком.
В задней части позвоночника позвонки видоизменяются: верхние и нижние дуги превращены в расширенные пластинки, поддерживающие наружные лучи хвостового плавника (рис. 49). Измененные нижние дуги называют гипуралиями (hypuralia), верхние –
 Рисунок 49 – Скелет хвостового плавника костистой рыбы:
Рисунок 49 – Скелет хвостового плавника костистой рыбы:
1 – гипуралии; 2 – уроневралии; 3 – уростиль.
уроневралиями (uroneuralia). Тела последних позвонков слиты в палочковидную косточку – уростиль (urostyl), направленную в верхнюю лопасть хвостового плавника. Гипуралии у некоторых рыб служат элементами, по которым определяют возраст.
В состав осевого скелета входят мускульные косточки (epipleuralia), примыкающие либо к невральным дугам, либо к телу позвонка, либо к ребрам.
У щуки общее число позвонков 56-65. Верхние концы дуг в туловищных позвонках не срастаются и образуют парные остистые отростки, но в заднем отделе тела, позади спинного плавника, верхние дуги, срастаясь, образуют непарные остистые отростки. Парапофизы у щуки двойные. Есть верхние и нижние межмышечные косточки.
У карпа число позвонков 36-38. Из скелетных элементов первых четырех позвонков образовалась система косточек, входящая в состав веберова аппарата, являющегося одной из отличительных особенностей карпообразных. Веберов аппарат – это орган, передающий изменение давления внешней среды во внутреннюю полость. Он представляет собой орган, состоящий из скелетных элементов первых позвонков, эндолимфатической полости лабиринта, перилимфатического пространства слуховой капсулы и плавательного пузыря (рис. 50). Остистый отросток первого позвонка преобразован в одну из косточек веберова аппарата – запор (claustrum); верхняя дуга первого позвонка – в чашевидную (scaphium) косточку. Из верхней дуги второго позвонка образована вставочная косточка (incus), из поперечного отростка третьего позвонка – трехногая (tripus). Последняя косточка веберова аппарата (os suspensorium) развивается за счет ребер четвертого позвонка. Все косточки связаны друг с другом, а трехногая с помощью связки соединена с наружной стенкой плавательного пузыря. При изменении наружного давления пассивно изменяется объем воздушной камеры, что влечет за собой движение косточек аппарата; в связи с этим изменяется давление через пери- и эндолимфу на лабиринт; передаваемое раздражение вызывает в качестве рефлекса соответствующее изменение мышечного тонуса. У карпа имеется большое число тонких косточек, располагающихся в толще мускулатуры и развивающихся за счет окостенения сухожилий.
Рисунок 50 – Веберов аппарат (вид сбоку):
1 – запор; 2 – чашевидная косточка; 3 – вставочная косточка; 4 – трехногая косточка; 5 – последняя косточка; 6 – ребра 5, 6 и 7-го позвонков; 7 – парапофизы 2-го и 4-го позвонков; 8 – верхние остистые отростки 2-7-го позвонков.
У налима общее число позвонков 58-66. Они несут ребра и парапофизы тонкие, длинные. Следует отметить, что у ряда тресковых рыб (навага, мерлуза) ребер нет, их функцию выполняют вздутия на парапофизах.
Скелет непарных плавников. Спинной и анальный плавники состоят из костных лучей: внутренних, скрытых в мускулатуре птеригофор (pterygiophora) и наружных плавниковых лучей – лепидотрихий (lepidotrichia) (рис. 51). У всех костистых рыб наблюдается соответствие числа птеригофор числу плавниковых лучей. Число лучей в плавниках и их характер является систематическим признаком.
Рисунок 51 – Скелет непарного спинного плавника костистой рыбы с прилегающим отделом позвоночника:
1 – лучи плавника – лепидотрихии (слева твердые, справа мягкие); 2 – птеригофоры.
Хвостовой плавник состоит из лепидотрихий, опирающихся на видоизмененные элементы последних позвонков – уростиль и гипуралии (см. рис. 49). У судака (окуня) хвостовой плавник гомоцеркальный (уростиль является границей между гипоуралиями и уроневралиями; большая часть лучей верхней лопасти и все лучи нижней лопасти опираются на гипуралии.
Скелет парных плавников и их поясов. Парные плавники состоят из поясов плавников и скелета свободного плавника. Опорой грудных плавников служит плечевой пояс. У судака (окуня) он представлен двумя хондральными и несколькими покровными костями (рис. 52). Хондральными являются: лопатка (scapula) и коракоид (coracoideum). Покровные кости: клейтрум (cleithrum), надклейтрум (supracleithrum), заднеключичная
Рисунок 52 – Плечевой пояс костистой рыбы с грудным плавником:
а – судака; б – карпа; 1 – ключица; 2 – надтеменная кость; 3 – задневисочная кость; 4 – надключичная кость; 5 – подключичная кость; 6 – лопатка; 7 – лучи плавника (лепидотрихии); 8 – радиалии; 9 – коракоид; 10 – мезокоракоид.
(postcleithrum), задневисочная (posttemporale) и надтеменная (supratemporale). Элементы первичного пояса неподвижно соединены с клейтрумом, а заднетеменные и надтеменные кости причленяются к осевому черепу, что обеспечивает прочную фиксацию плечевого пояса.
Скелет свободного плавника состоит из радиалий, отходящих от лопасти, а частично от коракоида, и лепидотрихий.
Тазовый пояс состоит из двух плоских треугольных костей (pubis), лежащих в толще брюшной мускулатуры (рис. 53). С осевым скелетом тазовый пояс не связан. У окунеобразных обычно тазовые кости прикреплены к ключицам. К боковым сторонам тазового пояса причленяются лучи брюшных плавников – лепидотрихий.
Рисунок 53 – Тазовый пояс костистой рыбы с брюшным плавником:
1 – тазовая кость; 2 – лучи плавника.
У щуки на месте задневисочной кости расположены две: верхняя называется задневисочной, а нижняя – надвисочной костью (supratemporale). Мезокоракоида нет. Колючих лучей в непарных и парных плавниках нет.
У карпа в плечевом поясе имеется дополнительная кость – мезокоракоид (mesocoracoideum). Впереди спинного плавника сохраняются многочисленные рудиментарные птеригофоры – остатки более длинного прежде спинного плавника. Лепидотрихии спинного и анального плавников в основном ветвистые. Первые 2-3 (4) луча неветвистые членистые, последние из них части бывают утолщены, теряют членистость, окостеневают и превращаются в шип, колючку, иногда сзади зазубренную.
У налима мезокоракоид в плечевом поясе отсутствует. Лопаточное отверстие расположено между лопаткой и коракоидом, тогда как у большинства рыб оно находится на самой лопаточной кости. В плавниках нет колючих лучей. Тазовый пояс соединен связкой с ключицами. Хвостовой плавник налима, как и других тресковых, дифицеркальный. Лучи верхней лопасти хвоста опираются на уроневралии, нижней – на гипуралии.
Рекомендации к разбору черепа судака (окуня). Разбор черепа рекомендуется начинать с висцерального скелета, снимая кости только с одной стороны головы.
1. Прежде всего следует снять мелкие окологлазничные косточки – элемент глазничного отдела осевого черепа – и после просушки разложить их в такой же последовательности, как они располагались на голове рыбы.
2. Снять с черепной коробки жаберную крышку. Самая большая кость – крышка; под ней лежит подкрышка; перед крышкой – изогнутая под углом зубчатая предкрышка, под ней расположена межкрышка. В верхнем углу крышечной кости (operculum) имеется углубление, через которое крышка соединяется с задним верхним концом гиомандибуляре.
3. Отделить кости вторичной верхней челюсти – предчелюстную и верхнечелюстную.
4. Удалив глаз и мускулатуру щеки, внимательно рассмотреть расположение и соединение костей. Впереди видна небная кость с зубами; связанная с ней изогнутая наружная крыловидная и лежащая между ними сверху в виде пластинки внутренняя крыловидная. К наружной крыловидной кости сзади примыкает округлая задняя крыловидная кость. Внизу она сочленяется с квадратной костью, имеющей треугольную форму. Квадратная кость присоединяется к нижней челюсти через сочленовную кость, сверху квадратная кость через симплектикум соединяется с гиомандибуляре.
5. Рассмотрев расположение костей челюстной дуги, следует перейти к вычленению отдельных элементов. Осторожно снять маленькую угловую кость; раздвинуть соединенные между собой зубную и сочленовные кости. Между ними обнаруживаются остатки меккелева хряща. Затем следует перейти к выделению костей небно-крыловидного отдела (не трогая подвесочную кость – гиомандибуляре) и после обработки разложить все кости, как показано на рис. 54.
Рисунок 54 – Схема расположение костей висцерального скелета головы судака:
I – челюстная дуга; II – гиоидная дуга; III – жаберная крышка; IV – носовая и окологлазничные кости мозгового черепа.
6. Нужно внимательно рассмотреть подъязычную дугу. Гиомандибуляре палочковидной костью соединяется со сложным гиоидом через верхнеподъязычную кость. Удаляем гиомандибуляре и палочковидную кость. Отчленив гиоид, начинаем его разбор. Впереди располагается непарная удлиненная кость – основная подъязычная. За ней следуют две лежащие друг над другом нижнеподъязычные кости. К ним снизу примыкает лежащая в толще мускулатуры непарная заднеподъязычная, или горловая, кость, а сверху – среднеподъязычная и верхнаеподъязычная кости. От последних снизу отходят тонкие изогнутые кости – лучи жаберной перепонки.
Все кости промываются, высушиваются, раскладываются в соответствии с рисунком и подклеиваются.
7. Из жаберных дуг вычленяем лишь первую жаберную дугу. Сверху вниз выделяем глоточно-жаберную, верхнежаберную, среднежаберную, нижнежаберную и непарную основную жаберную косточку, или копулу. Остальные жаберные дуги рекомендуется рассматривать по готовым макетам и рисункам.
8. Прежде чем перейти к разбору черепной коробки, необходимо рассмотреть на ней общее расположение видимых костей, руководствуясь рис. 55.
Рисунок 55 – Схема расположения костей черепной коробки:
I – обонятельный отдел; II – слуховой отдел; III – кости орбиты глаза; IV – затылочный отдел; V – крыша черепа; VI – дно черепа.
9. Разбор черепной коробки рекомендуется начинать с обонятельного отдела. Захватив пальцами переднюю часть черепной коробки, вытягиваем ее на себя. В этом отделе сохраняется много хряща, и кости необходимо отделить от него. Снизу хорошо отчленяется кость дна черепа – непарный сошник. Сверху лежит непарная средняя обонятельная кость, по бокам – парные боковые обонятельные кости. Отмыв и просушив кости, их следует разложить и наклеить на ватман в соответствии с рис. 55. Таким же образом надо поступать с костями остальных отделов после их разбора.
10. С нижней части черепной коробки надо удалить вторую кость дна черепа – длинный мечевидный парасфеноид.
11. После этого следует перейти к вычленению костей крыши черепа. Носовые кости снимаются при разборе висцерального скелета. Впереди лежат крупные, тесно соприкасающиеся друг с другом парные лобные кости; за ними – небольшие теменные, разделенные верхнезатылочной костью.
12. Разбирая глазничный отдел, надо отделить парные боковые клиновидные кости и непарную основную клиновидную, лежащую основанием на парасфеноиде и раздвоенными концами упирающуюся в основание переднеушной кости.
13. В слуховом отделе следует отчленить лежащие сверху клиновидноушные кости, затем крыловидно- и верхнеушные кости, предварительно найти отолиты, расположенные в области сочленения переднеушных костей с основной затылочной. В нижней передней части слухового отдела выделяем переднеушные кости. На крыловидно- и переднеушной костях в виде чешуйки лежит маленькая плоская заднеушная кость.
14. В затылочном отделе следует сначала отделить непарную верхнезатылочную кость, затем две боковые затылочные и основную затылочную кости.
Текст книги «Эволюционно-функциональная морфология животных»
Эти отличия дают основание для деления всех позвоночных на два раздела: Бесчелюстные (Agnatha), к которым относятся ныне живущие круглоротые и два вымерших класса, и Челюстноротые (Gnathostomata), включающие всех остальных позвоночных. Такая неравномерность объясняется тем, что активное участие висцерального аппарата в актах питания и дыхания давало челюстноротым несомненное преимущество в прогрессивной эволюции перед придонными или плавающими микрофагами-фильтраторами с неподвижной ротовой воронкой и малоактивными дыхательными движениями, какими были ныне вымершие классы бесчелюстных. В эволюционном «соревновании» с более активными челюстноротыми смогли сохраниться лишь формы, использовавшие сосущий тип ротового аппарата для специфического способа питания: круглоротые (миноги и миксины) присасываются предротовой воронкой к телу рыб и, разрушая роговыми «зубами» языка покровы жертвы, поршневидными движениями языка высасывают жидкость тканей и кровь. Миксины при этом глубоко вгрызаются в тело жертвы, перетирая с помощью роговых «зубов» языка все мягкие ткани.
Учитывая указанные принципиальные отличия, рассмотрим строение висцерального скелета круглоротых и рыб. У круглоротых (рисунок 12) он представлен хрящами, поддерживающими ротовую воронку и язык, а также ажурной хрящевой жаберной решеткой, поддерживающей стенки тела в месте расположения жаберных отверстий. Один из непарных хрящей предротовой воронки (задний верхний хрящ) прирастает к мозговой коробке; это единственная структура, связывающая висцеральный и осевой отделы черепа.
Челюстноротые, как уже говорилось, имеют висцеральный череп, состоящий из серии висцеральных дуг, расположенных в межжаберных перегородках. Передняя пара их функционирует в качестве подвижных челюстей. У хрящевых рыб (рисунок 13) челюстная дуга представлена двумя хрящами: нёбноквадратным, играющим роль верхней челюсти, и меккелевым, образующим нижнюю челюсть. Челюсти вооружены коническими, загнутыми назад зубами, происходящими от покрывающей тело плакоидной чешуи и состоящими из дентина, одетого снаружи чехликом из прочной эмали.
Следующая висцеральная дуга – подъязычная – включает два парных хряща: подвесок (hyomandibulare) и гиоид (hyoideum); гиоиды правой и левой подъязычных дуг объединяются непарным хрящом–копулой. Функция подъязычной дуги – подвижное сочленение висцерального аппарата к осевому черепу: подвесок своим верхним концом с помощью сустава соединен со слуховым отделом мозговой коробки, а нижним – с обоими элементами челюсти в области челюстного сустава. Такой тип связи висцерального и осевого отделов черепа называется гиостилией (рисунок 20) и характерен для большинства хрящевых рыб. У некоторых примитивных акул имеется дополнительная связь в виде отростка в передней части небноквадратного хряща, причленяющегося к дну мозгового черепа. Такой тип связи носит название амфистилии. У химер (подкласс Holocephali) верхняя челюсть полностью срастается с дном осевого черепа – возникает аутостилия.

I – амфистилический череп древней акулы; II – амфистилический череп костного ганоила; III – гиостилический череп современной акулы; IV – аутостилический череп наземного позвоночного (лягушка); / – нёбноквадратный хрящ; 2, 3 – выросты нёбноквадратного хряща; 4 – меккелев хрящ; 5 – нижняя челюсть; 6 – гиомандибуляре; 7 – гиоид.
Рисунок 20 – Типы причленения висцеральных дуг к мозговому черепу
Подъязычная дуга ограничивает сзади остаток жаберной щели, которая сохраняется у хрящевых рыб в виде брызгальца, соединяющего, как и все жаберные щели, полость глотки с внешней средой. У скатов, жаберные щели смещены на брюшную сторону, расположенное на спинной стороне брызгальце, играет важную роль в дыхании.
Позади подъязычной дуги расположены однотипно устроенные жаберные дуги. У большинства хрящевых рыб их пять – соответственно числу жаберных щелей: первая расположена позади подъязычной дуги, вторая – позади 1-й жаберной и т. д. У некоторых акул число жаберных шелей может достигать 6 – 7. Каждая жаберная дуга состоит из четырех подвижно сочлененных парных элементов, объединенных снизу непарной копулой. На жаберных дугах, кроме последней, расположены органы дыхания – жабры.
У костных рыб (рисунок 14) сохраняется тот же принцип строения висцерального черепа, но висцеральные дуги составлены не хрящевой, а костной тканью. В челюстной дуге образуется большое число первичных и вторичных костей. В верхней челюсти первичные кости представлены задней крыловидной и квадратной, а в нижней – сочленованной; квадратная и сочленованная кости образуют челюстной сустав. Вторичные кости в верхней челюсти представлены предчелюстной, челюстной, небной и двумя крыловидными, а в нижней – большой зубной костью и маленькой угловой. Предчелюстная, челюстная и зубная кости формируют вторичные челюсти, снабженные зубами; кроме того, зубы имеются на небной кости.
Череп костных рыб – типично гиостилический; соединение висцерального черепа с осевым осуществляется с помощью подъязычной дуги. Эта дуга образована первичными костями, соответствующими ее отделам у хрящевых рыб: гиомандибуляре (подвесок), гиоид и непарная копула. Кроме того, к подвеску прикрепляются кости, образующие жаберную крышку, – новое по сравнению с хрящевыми рыбами образование, обеспечивающее интенсификацию дыхательных движений. Эти кости – вторичные по происхождению.
Жаберные дуги (их у костистых рыб четыре) построены по тому же плану, что и у хрящевых рыб; их элементы составлены первичными костями. Пятая жаберная дуга редуцирована и не несет жабр.
Висцеральный череп амфибий в сравнении с рыбами изменен значительно. Личинки амфибий (и некоторые взрослые) имеют жабры, соответственно у них развиваются жаберные дуги. Количество жаберных дуг уменьшается до четырех; часто они не имеют расчленения на элементы и являются цельными.
У амфибий, прошедших метаморфоз, висцеральные дуги преобразуются в скелетные элементы, характерные для наземных позвоночных. Они входят в состав подъязычного аппарата, служащего опорой языку, в состав скелета гортани и кольцевидных хрящей трахеи, а также участвуют в совершенствовании слухового аппарата. Подъязычный аппарат располагается у лягушки в дне ротовой полости между ветвями нижней челюсти и представляет собой хрящевую пластину с рожками. Ее передние рожки, загибаясь вверх, охватывают кишечную трубку и прикрепляются к слуховым капсулам.
Функцию верхних челюстей у земноводных выполняет, как и у костистых рыб, костная дуга, состоящая из покровных межчелюстных (предчелюстных), верхнечелюстных и квадратных костей.
Элементы первичного челюстного комплекса, образованного исбноквадратным хрящом, принимают участие в формировании та мозгового черепа: нёбная кость прилегает к сошнику, а квадратная кость – к чешуйчатой кости. В нижней челюсти меккелев хрящ прикрыт снаружи зубной и угловой костями.
Череп амфибий аутостилический: челюстная дуга прирастает непосредственно к мозговому черепу (рисунок 15). Подъязычная дуга не принимает участия в прикреплении челюстного аппарата к черепу. Верхний элемент этой дуги – гиомандибуляре – прекращен в маленькую кость – стремя, которая, как подвесок у рыб, верхним концом упирается в слуховую капсулу. Она лежит в преобразованной из брызгальца полости и выполняет роль слуховой косточки.
Висцеральный скелет рептилий (ящериц) в значительной степени близок к амфибиям. Верхние челюсти образованы межчелюстными и верхнечелюстными костями, несущими зубы.
Нёбноквадратный хрящ подвергается дальнейшим превращениям. Хондральная квадратная кость своим верхним отделом связана с мозговой коробкой; снизу к ней причленяется нижняя челюсть. Остальная часть нёбноквадратного хряща покрывается кожными костями – в передней части нёбными, а в серединной – крыловидными. Все эти кости участвуют в укреплении дна черепа. Появляются новые покровные поперечные кости, соединяющие крыловидные кости с верхнечелюстными, а у ящериц и гаттерий еще и хондральные столбчатые кости, соединяющие крыловидные кости с теменными.
Нижняя челюсть, гомологичная меккелеву хрящу, состоит из хондральной сочленовной кости, которая причленяется к квадратной, и ряда накладных костей, облегающих меккелев хрящ: зубной, угловой, надугловой, венечной, пластинчатой. Верхний отдел подъязычной дуги, гомологичный гиомандибуляре, представлен (как и у земноводных) слуховой косточкой – стременем. Остальные элементы висцерального скелета образуют подъязычный аппарат, состоящий из хрящевой пластинки (гомолог копулы) и трех пар рожков (гомологи гиоидов и жаберных дуг).
Весьма своеобразно строение черепа у змей. Для них характерно подвижное соединение элементов челюстного аппарата и связанных с ними костей: нёбных, крыловидных и чешуйчатых. Эта особенность позволяет змее заглатывать пищу целиком, даже довольно крупных животных. Механизм заглатывания достигается попеременным движением левой и правой половин нижней челюсти. При наблюдении за кормящейся змеей создается впечатление, что она как бы наползает на свою добычу. У ядовитых змей подвижность костей облегчает перемещение ядовитых зубов (рисунок 21).

1 – ядовитый зуб; 2 – верхнечелюстная кость; 3 – поперечная ко сть; 4 – крыловидная кость; 5 – чешуйчатая кость; 6 – квадратная кость.
Рисунок 21 – Череп гремучей змеи с закрытой (А) и раскрытой (Б) пастью
У крокодилов зубы расположены на межчелюстных, верхнечелюстных и зубных костях и сидят в альвеолах, как у млекопитающих. В связи с разрастанием нёбных отростков межчелюстных и верхнечелюстных, а также нёбных костей у крокодилов развивается костный помост – вторичное костное нёбо. Оно разделяет полость рта на верхний отдел, носоглоточный ход, и нижний отдел, собственно ротовую полость (рисунок 22). Таким образом, внутренние отверстия ноздрей (хоаны) открываются не непосредственно в ротовую полость, а в ее заднюю часть, поэтому пища, находящаяся во рту, не мешает поступлению воздуха к органам дыхания. У черепах нёбо прирастает к мозговому черепу. Квадратная кость массивная и на большом протяжении контактирует с ушным отделом, повышая прочность задней части черепа.
В висцеральном черепе птиц много общего с черепом рептилий. Верхняя челюсть включает парные межчелюстные и верхнечелюстные кости. К последним причленяются палочковидные скуловые и квадратноскуловые кости, которые, в свою очередь, соединяются с квадратными костями. В итоге образуется весьма характерная для птиц нижняя височная дуга, отграничивающая снизу глазницу и височную яму. Нижняя челюсть состоит из гомологичной меккелеву хрящу сочленовной кости и из кожного происхождения зубной, пластинчатой, угловой и венечной костей. Вместе с верхней челюстью она образует характерный для птиц клюв. Челюсти, как правило, сильно вытянуты, их кости срастаются между собой. Возникает весьма совершенный, прочный хватательный аппарат (рисунок 18). Слуховая косточка, стремечко, одна, как и у рептилий.

1 – межчелюстные кости; 2 – верхнечелюстные кости; 3 – нёбные кости; 4 – крыловидные кости; 5 – хоаны.
Рисунок 22 – Вторичное костное у нёбо у крокодила
Костный подъязычный аппарат имеет вид удлиненной пластинки с очень длинными рожками, гомологичными первой паре жаберных дуг. Подъязычная пластинка участвует у птиц в движении языка при захвате пищи, а также при звукообразовании. Специалисты отмечают, что, например, у токующего глухаря во время второго колена призывной песни, когда птица глубоко втягивает язык в гортань, загнутые рожки сдвигают набухшую кровью кожу и соединительнотканную пленку на голове, отчего глухарь временно глохнет (рисунок 23).
Строение висцерального черепа млекопитающих в сравнении с рептилиями существенно меняется. Верхние челюсти состоят из парных межчелюстных и верхнечелюстных костей. Для всех млекопитающих характерно развитие вторичного костного нёба, образованного нёбными отростками межчелюстных и верхнечелюстных костей и нёбными костями. В связи с его образованием хоаны открываются не между верхнечелюстными костями, как у других наземных позвоночных, а как у крокодилов и черепах – позади нёбных костей. Такое строение нёба предотвращает закупорку хоан (т.е. перерыв дыхания) в то время, когда пищевой комок задерживается в ротовой полости при обработке пищи во рту и при глотании. Крыловидные кости уменьшаются и прирастают к дну черепа.

1 – язык; 2 – ороговевшие шипики языка; 3 – костная основа языка; 4 – передние рожки; 5 – задние рожки; а – обычное положение языка; б – язык втянут в глотку.
Рисунок 23 – Подъязычный аппарат глухаря (А) и его положение относительно слухового прохода (Б)
Нижняя челюсть образована только парными зубными костями, которые несут два отростка – венечный и угловой. Нижние челюсти причленяются непосредственно к чешуйчатым костям. Наиболее серьезным преобразованием становится превращение сочленовной кости в слуховую косточку – молоточек, а квадратной – в наковальню. Вместе со стременем они лежат в полости среднего уха и участвуют в звукопередаче. Наружная стенка последней, а также часть наружного слухового прохода окружены барабанной костью, гомологичной, по-видимому, угловой кости нижней челюсти прочих позвоночных. Таким образом, у млекопитающих наблюдается дальнейшее превращение части висцерального аппарата в слуховой аппарат среднего уха. Остатки подъязычного аппарата располагаются у млекопитающих в нижней части глотки и входят в состав части гортани. Функциональное значение скелета подъязычного аппарата определяется его ролью в движениях языка, связанных с захватыванием, обработкой в ротовой полости и проглатыванием пищи, с движениями глотки и гортани во время глотания, при воспроизведении звуков и т.д.
4.1.3 Скелет парных конечностей и их поясов
Ногообразные конечности наземных позвоночных произошли из парных плавникообразных конечностей предковых форм, живших в водной среде. В примитиве за исходную форму конечностей берется кожно-мышечная складка, которая проходила вдоль тела животного и выполняла роль рулей глубины и сохранения определенного положения тела в водном пространстве. Впоследствии средняя часть боковой складки редуцируется и образуются парные грудные и брюшные плавники, распологающиеся в менее подвижных участках осевого отдела туловища. Плавникообразные конечности, присущие всем рыбам, сохраняют первоначальную основную функцию складок, но приобретают большие возможности для адаптивных изменений, связанных как с совершенствованием локомоторной функции, так и с выполнением каких-либо дополнительных приспособлений.
У хрящевых рыб плавники располагаются в горизонтальной плоскости, выполняют роль рулей глубины и незначительную опорную функцию. Увеличение размеров плавников приводит к усилению в них хрящевой основы, которая первоначально представлена метамерно расположенными лучами. Хрящевые лучи своими основаниями, образуют общую хрящевую пластинку, которая затем преобразуется в скелет пояса конечности (рисунок 24).

А – плечевой пояс и грудные плавники: 1 – коракоидный отдел; 2 – лопаточный отдел; 3, 4, 5 – базалии; 6 – радиалий; 7 – эластоидиновые нити. Б – пояс задних конечностей и брюшные плавники: 1 – пояс; 2, 3, 4 – базалии; 5 – радиалий; 6 – эластоидиновые нити.
Рисунок 24 – Скелет поясов и парных конечностей самца акулы
У костистых рыб, с их более разнообразными приспособлениями к условиям существования, плавники стали выполнять функцию изменения направления движения, роль органов опоры (особенно у двоякодышащих), и даже в осуществлении планирующего полета (летающие рыбы). Такие адаптации сопровождались усилением хрящевого пояса, охватывающего тела со всех сторон, и образованием на его боковой поверхности выступа для соединения с главным, или осевым, лучом плавника. Плавник по отношению к осевому отделу туловища стал занимать не горизонтальное, а наклонное или даже поперечное положение, в результате чего бывшая дорсальная поверхность становится краниолатеральной. Усиление функциональной нагрузки на плавникообразные конечности привело к дополнительному усилению их хрящевого остова костными образованиями кожного происхождения, которые затем постепенно замещают хрящевой скелет. Одновременно происходит и усложнение костной основы плавника, где наряду с расчленением конечностей на звенья происходит и их дифференциация. Брюшные плавники, имеющие меньшую функциональную загруженность, длительное время сохраняют и более примитивное строение (рисунок 25).
Первичный пояс: 1 – лопатка; 2 – коракоид. Вторичный пояс: 3 – клейтрум; 4 – супраклейтрум; 5 – заднетеменная кость; 6 – подклейтрум. Скелет плавника: 7 – радиалий; 8 – лепидотрихии (кожные костные лучи).
Рисунок 25 – Грудной пояс костистой рыбы судака
У наземных позвоночных за счет соединения вентральных концов нижниx ребер образуются парные хрящевые пластинки, которые, срастаясь между собой, образуют тело грудины. У амфибий в связи с вторичной редукцией нижних ребер грудина развивается независимо от ребер и имеет непосредственное отношение к плечевому поясу, так как служит местом прикрепления коракоидных костей. В связи со сменой среды обитания на воздушную плавникообразные конечности преобразуются в короткие ногообразные конечности с характерным подразделением их остова на скелет пояса и скелет свободного отдела, как и у последующих классов наземных позвоночных, подразделенного на три основных звена – плечо – бедро, предплечье – голень, кисть – стопа (рисунок 26). Несмотря на наличие коротких ногообразных конечностей, основной принцип движения для хвостатых амфибий, точно так же и для ящерищеобразных, сохраняется прежний – боковые изгибы осевого отдела туловища и хвоста.
А– передняя конечность; Б – задняя ко ценность; I – плечо – бедро; II – предпечье – голень; III – кисть – стопа; 1 – плечевая кость; 2 – локтевая кость; 3 – лучевая кость; 4 – запястье; 5 – пясть; б – фаланги пальцев; 7 – бедренная кость; 8 – большая берцовая кость; 9– малая берцовая кость; 10 – предплюсна; 11 – плюсна 12 – коленная чашечка.
Рисунок 26 – Схема строения парных конечностей наземных позвоночных
Скелет плечевого пояса у низших наземных позвоночных сохраняет многие черты такового костистых рыб. Однако в связи с большим значением при опоре и передвижении животного его отделы приобретают большую дифферецированность и усиление за счет развития аналогичных костных образований кожного происхождения, которые в конечном итоге преобразуются в дорсальный – лопатку и вентральный отделы – коракоид и ключицу (рисунок 27). Наибольшее развитие приобретают коракоидиая кость, как опорное звено, и лопатка, обеспечивающая прочное соединение конечности с туловищем.
Тазовый пояс, как и плечевой, имеет те же отделы, из которых большее развитие получил вентральный, представленный; седалищной и лонной костями, в то время как подвздошная, относящаяся к дорсальному отделу развита значительно слабее (рисунок 27). Она имеет вытянутую заостренную форму, направлена дорсокаудально и соединяется с боковой поверхностью ребер и одним из позвонков.
У рептилий, в частности у ящериц и крокодилов, плечевой пояс включает в основном те же элементы, что и у амфибии (рисунок 28).
Брюшная его часть представлена коракоидами, несущими сочленовные ямки для причленения плечевых костей. Спинная часть пояса состоит из лопатки и надлопаточного хряща. Коракоид связан с грудиной и обладает подвижностью. Впереди от него расположена ключица. Характерно наличие крестообразного надгруднаника, лежащего перед грудиной и связывающего ее с ключицами. В целом плечевой пояс рептилий более прочен, чем у амфибий, и эта его черта отражает их лучшую приспособленность к передвижению по суше.
В тазовом поясе парные кости: подвздошная, седалищная и лобковая соединены в единый замкнутый таз. Спереди и сзади, на стыке костных элементов расположены передний и задний хрящевые отростки. Подвздошными костями пояс причленяется к поперечным отросткам двух крестцовых позвонков. На стыке трех костей таза расположена вертлужная впадина: в нее входит круглая головка бедренной кости.
1 – грудина: 2 – хрящевая задняя часть грудины; 3 – коракоид; 4 – ключица лежит на прокоракоиде; 5 – предгрудина; 6 – лопатка; 7 – сочленовная впадина на лопатке для плеча (хрящ покрыт точками); 8 – подвздошная кость; 9 – седалищная кость; 10 – вертлужная впадина; 11 – крестцовый позвонок, 12 —уростиль.
Рисунок 27 – Плечевой (А) и тазовый (Б) пояс лягушки (вид спереди)
А: 1 – ключица; 2 – надлопаточный хрящ; 3 – лопатка; 4 – коракоид; 5 – ребра; 6 – грудина; 7 – переднекоракоидный хрящ; 8 – надгрудинник. Б: 1 – суставная впадина для головки бедра; 2 – подвздошная кость; 3 – лобковая кость; 4 – седалищная кость; 5 – связка; 6 – «окно»; 7 – запирательное отверстие; 8 – задний хрящевой отросток; 9 – передний хрящевой отросток.
Рисунок 28 – Плечевой (А) и тазовый (Б) пояс ящерицы
Передняя и задняя конечности имеют типичное строение. Особенность конструкции суставов в конечностях рептилий иная, чем у амфибий: сочленение в передней конечности лежит между костями запястья (межзапястный, или интеркарпальный, тип), а в заднем – между костями предплюсны (межпредплюсневый, или интертарзальный, тип). Конструкция сустава создает сгибательно-разгибательную подвижность конечности. Сходное строение имеют суставы птицы.
У птиц скелет передних конечностей (пояс и свободная конечность) имеет черты, преемственные от рептилий, но в связи с приспособлением к полету отличается рядом особенностей. Пояс включает лопатки, коракоиды (вороньи кости) и ключицы, которые своими проксимальными концами формируют площадки для причленения плечевых костей. Лопатка длинная, саблевидно изогнутая, лежит на ребрах, по которым она может свободно скользить. Коракоид развит очень хорошо. Одним концом он упирается в грудину, другой конец дает опору для причленения плеча. Грудина, являющаяся местом прикрепления мощной вентральной группы мышц плечевого пояса и выполняющая основную функциональную нагрузку при полете получает наиболее сильное развитие. Левая и правая ключицы срастаются между собой, образуют характерную для птиц вилочку, придающую поясу особую упругость.
Скелет крыла состоит из всех типичных для пятипалой конечности отделов (рисунок 29). При этом плечо и предплечье существенно не изменены, а кисть, наоборот, имеет ряд своеобразных особенностей. Сильно видоизменено запястье. Его проксимальные части срастаются в две косточки, а дистальные – с пястью. Пясть состоит из двух удлиненных костей, сращенных как в проксимальном, так и в дистальном отделах и образующих в итоге сложную пястно-запястную кость, или пряжку.
1 – кисть (первостепенные); 2 – предплечье (второстепенные); 3 – плечо (третьестепенные).
Рисунок 29 – Скелет крыла птицы и расположение маховых перьев
Из пальцев сохраняются только три – второй, третий и четвертый, при этом только третий палец имеет две фаланги, а второй и четвертый – по одной. В связи с наличием особых мускулов, сухожилий и кожистых перепонок кости крыла соединены между собой так, что все крыло не может быть вытянуто по прямой, и его части расположены под некоторым углом друг к другу. Отделы конечности могут двигаться только в одном направлении – в плоскости крыла, складывая и расправляя его.
Скелет задних конечностей (рисунок 30) (пояс и свободная конечность) птиц также имеет ряд особенностей, связанных с тем, что при движении по земле птица переносит на них всю тяжесть тела. Прочность осевого корпуса тела определяется срастанием больших подвздошных костей по всей их длине со сложным крестцом. С подвздошными костями срастаются также весьма крупные седалищные кости. Лобковые кости, наоборот, малы; в виде тонких палочек они присоединены к наружному краю костей седалищных. Все три кости таза участвуют в образовании вертлужной впадины. В отличие от других наземных позвоночных (рептилий, млекопитающих) у птиц отсутствует лобковый симфиз – сочленение тазовых костей внизу: две половины тазового пояса широко расставлены, что связано с откладыванием крупных яиц, покрытых твердой скорлупой, и необходимостью в широком тазовом выходном отверстии. Такой тип таза называют открытым. Закрытый таз имеют только африканские страусы.
Бедро типичного строения. Голень состоит из двух типичных костей, однако полностью развивается только большаяперцовая кость, малая берцовая кость рудиментарна и прирастает к первой. К дисталыюй части голени прирастает проксимальный ряд косточек предплюсны. Лежащий вслед за голенью отдел конечности носит название цевки. У взрослой тины он состоит из одной длинной кости, подвижный сустав у птиц (как и у пресмыкающихся) расположен между двумя рядами костей предплюсны, а не между предплюсной и голенью, как у амфибий. Это сочленение именуется межпредплюсневым, или интертарзальпым.
1 – бедро; 2 – большая берцовая кость; 3 – редуцироманная малая берцовая кость; 4 – цевка; 5 – чашечка.
Рисунок 30 – Скелет задней конечности голубя
Дальнейшие дифференциация и специализация грудных и тазовых конечностей наземных позвоночных стали возможными лишь после поворота конечностей из трансверзальной (поперечной) плоскости в боковую, отрыва тела животного от земли и подведения конечностей под туловище. Эти преобразования привели к усилению дорсальных отделов поясов и превращению конечностей в активные органы локомоции. На грудных конечностях в связи с усилением грудной кости и развитием лопатки произошла полная редукция коракоида, а с утратой элементов мультифункциональности – значительное ослабление или редукция ключицы. Редукция вентральных звеньев плечевого пояса привела к значительному усилению мышц, обеспечивающих укрепление тела животного междуконечностями в подвешенном состоянии.
Из-за того, что тазовые конечности стали выполнять роль основных движителей в осуществлении поступательного движения, в тазовое поясе произошли значительное усиление дорсального отдела, смещение его в дорзосокраниальное положение и прочное сращение с позвоночным столбом. Кости вентрального звена срослись между собой по вентральной срединной линии и приняли участие в образовании вентральной срединной полости, которая стала продолжением брюшной. Включение тазового пояса в состав осевого отдела привело к тому, что он стал основным передатчиком двигательных усилий, исходящих от тазовых конечностей. Свободные отделы грудных и тазовых конечностей, имеющих в примитиве аналогичные по строению и гомологичные по происхождению звенья, с перемещением конечностей из трансверзальной плоскости в парамедианную испытали характерные изменения в их взаимоотношения.
На грудных конечностях при подведении их под туловище при незначительной изменении положения костей кисти поворот отдельных звеньев осуществлялся в плечевом и локтевом суставах, что привело к Хобразному перекресту костей предплечья. Каудальная поверхность плеча стала медиальной, а вершина локтевого сустава, имевшая дорсолатеральное направление, стала располагаться с каудальной стороны конечности. На тазовых конечностях, в отличие от грудных, поворот осуществлялся всем свободным отделом так, что бывшая латеральная поверхность стала краниальной, а краниальная – медиальной. Кости стопы, имевшие латерокаудальное направление пальцев, стали располагаться краниально. В связи с одновременным перемещением всех звеньев конечностей Х-образного перекреста костей голени не произошло, и поэтому у всех видов животных малая берцовая кость на голени занимает латеральное положение. Дальнейшая специализация аппарата движения происходила в зависимости от характера выполняемых функций и типа опоры животного.
У животных с развитием и совершенствованием скорости передвижения в процессе эволюции происходит смена опоры от стопо– через пальце– к фалангохождению (рисунок 31).
При смене опоры изменения в строении конечностей касаются не только редукции боковых пальцев, как это имеет место у парнокопытных (жвачные, нежвачные, мозоленогие), и непарнокопытных (тапиры, носороги и лошади), но и всех выше расположенных звеньев, которые выразились в изменении их длины, толщины, рельефа суставных поверхностей, степени развития связок, соответствующих мышц сосудов и нервов.
А – обезьяна павиан; Б – собака; В – лама.
Рисунок 31 – Задние конечности пальце– и стопоходящих млекопитающих. Элементы стопы затемнены
Знание коррелятивных изменений в строении периферического скелета при различных адаптациях позволяет при анализе палеонтологических материалов вскрыть основные закономерности эволюции конкретных видов животных, как это было сделано В.О. Ковалевским в отношении современной лошади. Он убедительно доказал, что одним из древнейших предков современной лошади был фенакод, который жил в начале третичного периода (около 50 млн. лет назад) в восточной части Северной Америки, где условия того времени характеризовались теплым влажным климатом и мягкими почвами с богатой сочной растительностью. Фенакод имел небольшие размеры (68 см в высоту) и пятипалую конечность. Из пяти пальцев два боковых были самыми короткими, а из трех других средний имел наибольшую длину с роговым башмачком на конечной фаланге, что свидетельствует о ведущей роли при передвижении.
Таким образом, внутренний скелет хордовых энто– и мезодермального происхождения поддерживает тело, служит для укрепления скелетных мышц, участвует в движении, обеспечивает защиту внутренних органов. Прогрессивные изменения связаны с заменой в осевом скелете единого нечлененного упругого тяжа – хорды сначала хрящевым, а затем костным позвоночником, его последовательной дифференцировкой на отделы и появлением мозгового черепа. В висцеральном скелете в эволюционном ряду позвоночных висцеральные дуги редуцируются, развиваются челюсти, формируются слуховые косточки. Мозговой и висцеральный скелет объединяются в единое морфофункциональное образование – череп с лицевой и мозговой частями. Скелет органов движения у первичноводных хордовых – первоначально непарных плавников – прогрессивно развивается в связи со специализацией к условиям обитаний, затем дополняется парными плавниками, используемыми как стабилизаторы и рули при движении и поворотах. Появление мускулатуры на унисериальных парных плавниках, использовавшихся в качестве опоры при движении по грунту дна, послужило основой для формирования пятипалой конечности наземного типа.
Карташев Н.Н., Соколов В.Е., Шилов И.А.
Практикум по зоологии позвоночных
М., Высшая школа. 1981 г.
ТЕМА 7. СКЕЛЕТ КОСТИСТОЙ РЫБЫ
СИСТЕМАТИЧЕСКОЕ ПОЛОЖЕНИЕ ОБЪЕКТА
Подтип Позвоночные, Vertebrata
Класс Костные рыбы, Osteichthyes
Подкласс Лучеперые, Actinopterygii
Группа Костистые рыбы, Teleostei
Представитель — Судак, Lucioperca lucioperca L.
МАТЕРИАЛ И ОБОРУДОВАНИЕ
Смонтированный скелет судака на подставке — один на всю группу. На одного-двух студентов необходимы:
1. Смонтированный на картонных планшетах разборный скелет судака.
2. Препарировальные иглы — 2.
ЗАДАНИЕ
Рассмотреть скелет в целом. Найти его основные части: череп, позвоночник, скелет парных конечностей и их поясов, скелет непарных конечностей. Рассмотреть детали строения отдельных частей скелета. Сделать следующие рисунки:
1. Осевой череп сбоку.
2. Висцеральный череп сбоку.
3. Грудной пояс с плавником.
4. Тазовый пояс с плавником.
5. Туловищный позвонок спереди.
6. Хвостовой позвонок спереди.
7. Продольный разрез нескольких позвонков.
8. Хвостовой плавник.
ОПИСАНИЕ СКЕЛЕТА
Осевой скелет (позвоночник). Осевой скелет костистых рыб составлен многочисленными костными позвонками. Тела позвонков спереди и сзади вогнутые — такие позвонки называют амфицельными. Пространство, образующееся между вогнутыми поверхностями соседних позвонков, и узкий канал, пронизывающий в центре тела позвонков, заполнены остатками хорды (рис. 34, 1), имеющей четковидную форму. Позвоночник делится на два отдела: туловищный (pars thoracalis) и хвостовой (pars caudalis); позвонки этих отделов отличаются своим строением.
Рис. 34. Схема продольного разреза хвостового отдела позвоночника судака:
1 — редуцированная хорда, 2 — тело позвонка, 3 — верхняя дуга, 4 — верхний остистый отросток, 5 — канал для спинного мозга, 6 — нижняя дуга, 7 — нижний остистый отросток, 8 — гемальный канал
Рис. 35. Позвонки судака
А — туловищный позвонок; Б — хвостовой позвонок:
1 — тело позвонка, 2 — поперечный отросток, 3 — ребро, 4 — верхняя дуга, 5 — верхний остистый отросток, 6 — нижняя дуга, 7 — нижний остистый отросток, 8 — мышечная косточка
Туловищные позвонки (рис. 35, А) имеют округлое в поперечном сечении тело (corpus vertebrae, 35, 1), от которого в стороны отходят поперечные отростки (processus transversus, рис. 35, 2). К этим отросткам причленяются ребра (costa, рис. 35, 3), а к некоторым ребрам — тоненькие мышечные косточки (рис. 35, 8). От верхней (спинной) части позвонка отходят верхние дуги (arcus neuralis, рис. 35, 4), замыкающиеся остистым отростком (processus spinosus, рис. 35, 5). В канале, образованном верхними дугами позвонков, располагается спинной мозг.
Хвостовые позвонки (рис. 35, Б) также имеют тело и верхние дуги, но поперечные отростки их смещены вниз и образуют нижние дуги (arcus haemalis; рис. 35, 6), замыкающиеся непарным нижним остистым отростком (processus spinosus inferior; рис. 35, 7). Канал, образуемый нижними дугами хвостовых позвонков, носит название гемального; в нем располагаются хвостовые артерия и вена. Расположение этих крупных кровеносных сосудов внутри костного канала предохраняет их от сдавливания при сокращениях мощной мускулатуры хвоста.
Череп. Как и у хрящевых рыб, череп костистых рыб состоит из двух отделов: осевого черепа, или мозговой коробки (neurocranium), и лицевого, или висцерального, черепа (splanchnocranium). Но в отличие от хрящевых череп костистых рыб почти целиком образован костной тканью и состоит из многочисленных отдельных костей.
Рис. 36. Осевой (мозговой) череп судака.
А — сбоку; Б — сверху; В — снизу:
1 — основная затылочная кость, 2 — боковая затылочная кость, 3 — верхняя затылочная кость, 4 — ушные кости, 5 — глазница, 6 — боковая клиновидная кость, 7 — боковая обонятельная кость, 8 — средняя обонятельная кость, 9 — лобная кость, 10 — теменная кость, 11 — носовая кость, 12 — парасфеноид, 13 — сошник, 14 — основная клиновидная кость
Осевой череп костистых рыб подразделяется на те же отделы, что и у хрящевых рыб. В каждом отделе образуется несколько костей. Затылочный отдел черепа составлен четырьмя костями: основной, или нижней, затылочной (basioccipitale; рис. 36, 2; рис 37, 1). Двумя боковыми затылочными (occipitale laterale; рис. 36, 2, рис. 37, 2) и верхней затылочной (supraoccipitale; рис. 36, 3; рис. 37, 3) Эти кости окаймляют большое затылочное отверстие (foramen occipitale magnum; рис. 37, 3), через которое головной мозг соединяется со спинным.
Рис. 37. Затылочный отдел осевого черепа судака:
1 — основная затылочная кость, 2 — боковая затылочная кость, 3 — верхняя затылочная кость, 4 — затылочное отверстие, 5 — ушные кости
В слуховом отделе черепа располагается по пять ушных костей (ossa otici; рис. 36, 4) с каждой стороны. В глазничном отделе имеются парные боковые клиновидные (laterosphenoideum; рис. 36, 6) и глазоклиновидные (orbitosphenoideum) кости. Последние расположены на тонкой межглазничной перегородке и при препарировке, как правило разрушаются; у судака их нет. В основании мозговой коробки образуется основная клиновидная кость (basisphenoideum); при рассматривании черепа сбоку виден лишь ее передний конец (рис. 36, А, 14). Впереди, в области обонятельного отдела, находятся кости парные боковые обонятельные (ectoethmoideum, рис. 36, 7) и непарная средняя обонятельная (mesethmoideum, рис. 36, 8). Все эти кости по происхождению замещающие (первичные), они возникли путем замещения соответствующих участков хрящевого черепа костной тканью.
Кости, прикрывающие череп сверху и снизу, по происхождению относятся к покровным (вторичным) костям. Они закладываются в соединительно-тканном слое кожи и позже, погружаясь под кожу, прирастают к хрящевой основе черепа. Среди них в крыше черепа бросаются в глаза крупные плоские лобные кости (frontale; рис. 36, 9), покрывающие большую часть черепной коробки. Сзади от них по обе стороны от гребня верхней затылочной располагаются относительно небольшие теменные кости (parietale; рис. 36, 10). Спереди от лобных в области обонятельного отдела находятся парные носовые кости (nasalе; рис. 36, 11), разделенные уже упоминавшейся средней обонятельной костью.
Дно черепа прикрыто крупной непарной костью, называемой парасфеноидом (parasphenoideum; рис. 36, 12). Впереди парасфеноида расположен непарный сошник (vomer; рис. 36, 13), в своей передней части снабженный зубами.
Рис. 38. Череп судака сбоку:
1 — окологлазничные кости, 2— слезная кость, 3 — нёбная кость, 4 — квадратная кость, 5 — крыловидные кости, 6 — сочленовная кость, 7 — предчелюстная кость, 8 — верхнечелюстная кость, 9 — зубная кость, 10 — угловая кость, 11 — подвесок, 12 — симплектикум, 13 — гиоид, 14 — копулагиоида, 15 — крышечная кость, 16 — предкрышечная кость, 17 — подкрышечная кость, 18 — межкрышечная кость
При рассматривании черепа сбоку помимо уже отмеченных костей видно полукольцо из мелких косточек, окаймляющих глазницу — окологлазничных (ossa infraorbitalia; рис. 38, 1). Передняя из них носит особое название — слезная кость (lacrimale; рис. 38, 2). Все эти кости покровные.
Висцеральный череп костистых рыб представлен серией висцеральных дуг: челюстной (самая передняя), подъязычной и пятью жаберными, из которых последняя (задняя) заметно редуцирована.
В челюстной дуге можно различить первичные и вторичные челюсти. Первичные челюсти образованы преимущественно замещающими костями и гомологичны хрящам нёбно-квадратному и меккелеву хрящевых рыб. В верхней челюсти они представлены спереди смешанной нёбной костью (palatinum; рис. 38, 3), сзади (в месте сочленения с нижней челюстью) — квадратной костью (quadratum; рис. 38, 4); между ними расположены три крыловидные кости (pterygoideum; рис. 33, 5). Из них лишь одна (задняя крыловидная — metapterygoicieum) замещающая, тогда как остальные две — наружная крыловидная (ectopterygoideum) и внутренняя крыловидная (entopterygoideum) — по происхождению покровные.
В нижней челюсти первична лишь одна сочленовная кость (articulare; рис. 38, 6), осуществляющая подвижное причленение нижней челюсти к верхней (через квадратную кость).
Вторичные челюсти представлены предчелюстными (praemaxillare; рис. 38, 7) и верхнечелюстными (maxillare; рис. 38, 8) костями в верхней половине и крупной зубной костью (dentale, рис. 38, 9) — в нижней. Кроме того, в нижней челюсти имеется еще маленькая угловая кость (angulare; рис. 38, 10), приросшая к задне-нижнему краю сочленовной. Предчелюстная и зубная кости снабжены довольно крупными зубами. Вторичным челюстям принадлежит главная хватательная функция. Дополнительную функцию удерживания добычи выполняют зубы, расположенные на нёбных костях и сошнике.
Все перечисленные кости челюстной дуги — парные; они симметрично повторяются на правой и левой половинах челюстей.
Подъязычная дуга целиком составлена первичными (замещающими) костями. Верхний элемент ее — подвесок (hyomandibulare, рис. 38, 11) верхним концом причленяется к слуховому отделу осевого черепа, а нижним — через маленькую дополнительную косточку (symplecticum; рис. 38, 12) — к квадратной кости челюстной дуги. Кроме этого, подвесок срастается с задней крыловидной костью. Таким образом, подвесок выполняет функцию причленения челюстной дуги к осевому черепу; иными словами, череп у костистых рыб, как и у акуловых, гиостилический. Нижняя ветвь подъязычной дуги, гомологичная гиоидному хрящу акуловых рыб, представлена несколькими окостенениями. Из них крупный — гиоид (hyoideum; рис. 38, 13). Гиоиды правой и левой сторон соединяются через непарную кость (copula; рис. 38, 14), одновременно поддерживающую язык. К гиоиду прикрепляются тонкие изогнутые косточки — лучи жаберной перепонки, поддерживающие кожистый край жаберной крышки.
Жаберные дуги представлены каждая четырьмя парными косточками, подвижно сочлененными между собой и объединяющимися снизу при помощи непарных костных элементов — копул. Исключение составляет лишь пятая, самая задняя жаберная дуга, имеющая лишь один парный (нижний) отдел.
Жаберные крышки являются по сравнению с хрящевыми рыбами новообразованием. Каждая состоит из четырех вторичных (покровных) костей: крышки (operculum; рис. 38, 15), предкрышки (praeoperculum; рис. 38, 16), подкрышки (suboperculum; рис. 38, 17) и межкрышки (interoperculum; рис. 38, 18). Жаберная крышка каждой стороны через предкрышечную кость прикрепляется к соответствующему подвеску и к квадратной кости.
Парные конечности и их пояса. Парные конечности представлены грудными и брюшными плавниками. Опорой грудных плавников в теле рыбы служит плечевой пояс (рис. 39). Он представлен двумя небольшими замещающими (первичными) и несколькими покровными костями. Верхняя из замещающих костей — лопатка (scapula; рис. 39, 1) — расположена в области причленения свободной конечности (ее легко отличить по небольшому круглому отверстию в центре кости). Сразу под ней находится врановая кость, или коракоид (coracoideum; рис. 39, 2). Эти два элемента составляют первичный пояс. Они неподвижно соединены с крупной покровной костью клейтрум (cleithrum; рис. 39, 3), верхний конец которой направлен несколько вперед; к нему присоединяется небольшая кость надклейтрум (supracleithrum; рис. 39, 4).
Рис. 39. Плечевой пояс и грудной плавник судака:
1 — лопатка, 2 — коракоид, 3 — клейтрум, 4 — надклейтрум, 5 — заднетеменная кость, 6 — заднеключичная кость, 7 — радиалии, 8 — костные кожные лучи
Клейтрум в свою очередь соединяется с заднетеменной (posttemporale; рис. 39, 5) костью. Направленные вперед нижние концы правого и левого клейтрумов соединяются друг с другом. Позади клейтрума неподалеку от лопатки и коракоида расположена небольшая заднеключичная кость (postcleithrum; рис. 39, 6). Все названные кости парные; они составляют вторичный плечевой пояс. Правая и левая заднетеменные кости причленяются к осевому черепу, что обеспечивает более прочную фиксацию пояса и таким образом усиливает его опорную функцию.
Грудной плавник в своем основании имеет один ряд мелких косточек — радиалии (radialia; рис. 39, 7), отходящих от лопатки (частично и от коракоида). Вся свободная лопасть плавника состоит из членистых кожных лучей1 (lepidotrichia; рис. 39, 8). Особенность скелета грудных плавников костистых рыб, по сравнению с хрящевыми, заключается в редукции базалий.
Подвижность грудных плавников увеличивается потому, что мышцы прикрепляются к расширенным основаниям кожных лучей, подвижно сочленяющихся с радиалями.
Рис. 40. Тазовый пояс и брюшной плавник судака:
1 — тазовые кости, 2 — кожные костные лучи
Тазовый пояс (рис. 40) представлен сливающимися друг с другом парными плоскими треугольными костями, лежащими в толще брюшной мускулатуры и не связанными с осевым скелетом. К боковым сторонам тазового пояса причленяются брюшные плавники. У большинства костистых рыб в скелете брюшных плавников отсутствуют базалий и полностью редуцировались радиалии: лопасть плавника поддерживается кожными костными лучами (lepidotrichia), расширенные основания которых непосредственно причленяются к тазовому поясу. Такое упрощение скелета брюшных плавников, видимо, связано с их ограниченными функциями.
Непарные конечности. Непарные конечности представлены спинными, подхвостовым (анальным) и хвостовым плавниками. Анальный и спинные плавники состоят из костных лучей, подразделяющихся на внутренние (скрытые в толще мускулатуры) птеригофоры и наружные плавниковые лучи — лепидотрихии.
Рис. 41. Хвостовой плавник судака:
1 — конец позвоночного столба — уростиль, 2 — гипуралин, 3 — костные кожные лучи — лепидотрихии
Хвостовой плавник, как это было видно уже при наружном осмотре, имеет внешне равнолопастное строение, однако при рассмотрении его внутреннего скелета (рис. 41) видно, что концевые позвонки позвоночного столба сливаются в палочковидную косточку — уростиль (urostyl; рис. 41, 1), которая заходит в основание лишь верхней лопасти плавника, а основание нижней лопасти поддерживается разросшимися, довольно широкими нижними дугами позвонков — гипуралиями (рис. 41, 2). Такой тип строения хвостового плавника, вполне или почти симметричного по внешней форме, но асимметричного по расположению внутреннего скелета, носит название гомоцеркального. Наружный скелет хвостового плавника составлен многочисленными кожными лучами — лепидотрихиями (рис. 41, 3).
ЗАКЛЮЧЕНИЕ
Общий тип организации водных челюстноротых позвоночных, сформировавшийся у хрящевых рыб, претерпевает дальнейшее развитие в классе костных рыб. Сохраняя общий план строения предков, костные рыбы в своей эволюции приобрели ряд прогрессивных черт, способствовавших повышению уровня жизнедеятельности и широкой адаптивной радиации этих животных.
Наиболее существенное отличие костных рыб от хрящевых заключается в развитии у них костного скелета. Биологическое значение этого явления очень велико: превосходя хрящ в прочности, костная ткань придает скелету еще и большую легкость и подвижность.
Развитие костной ткани происходит двумя независимыми путями. С одной стороны, имеют место довольно сложные гистологические процессы, в результате которых хрящевая ткань, составляющая основу скелета зародыша, постепенно замещается костной. Так возникают замещающие (хрящевые, или первичные) кости. Первичный костный скелет принимает на себя практически все те функции, которые у хрящевых рыб принадлежали хрящевому скелету.
С другой стороны, костная ткань может формироваться непосредственно в соединительнотканном слое кожи независимо от хряща. Таким путем развиваются покровные (кожные, или вторичные) кости. Именно так возник в эволюции костных рыб ряд новых элементов скелета, имеющих большое биологическое значение. Не будучи связанными с первичными (исходными) функциями хрящевого скелета, вторичные костные образования открывают возможность большого разнообразия форм, отражающих приспособления к разным условиям жизни. Таковы, например, вторичные челюсти, принимающие на себя хватательную функцию и отличающиеся большим разнообразием строения у разных видов в связи с приспособлением их к различным типам питания, гораздо более многообразным, чем у хрящевых рыб.
Только вторичные кости образуют жаберную крышку, имеющую большое значение в активизации дыхания. Вторичный плечевой пояс осуществляет связь первичного пояса с черепом. При характерном для рыб неподвижном соединении черепа с позвоночником такая связь определяет собой более прочную фиксацию пояса и усиление его опорной функции.
В эволюции рыб сначала покровные кости возникали в добавление к хрящевому скелету (состояние, характерное, например, для осетровых рыб), а позднее начался процесс окостенения хряща и образование замещающих костей; одновременно покровные окостенения начали погружаться под кожу, соединяясь с замещающими (хрящевыми) костями.
Во внутреннем строении костистых рыб наиболее бросающаяся в глаза особенность — появление плавательного пузыря — гидростатического органа, увеличивающего «плавучесть» и позволяющего рыбам маневрировать без существенной затраты энергии. У хрящевых рыб это возможно только при движении, что, естественно, требует значительной затраты энергии. Плавательный пузырь выполняет и некоторые добавочные функции: служит резонатором издаваемых рыбой звуков, может служить резервуаром для накопления резервного запаса кислорода (а у некоторых видов — и органом воздушного дыхания) и т. п.
Отсутствие спирального клапана, свойственного хрящевым рыбам, компенсируется у костистых рыб увеличением относительной длины кишечника и развитием у многих видов пилорических придатков, также увеличивающих общую всасывающую поверхность кишечника. Эти преобразования способствуют усилению интенсивности и эффективности пищеварения.
Своеобразно строение мочеполовой системы костистых рыб. Они, как и хрящевые рыбы, обладают мезонефрическими (туловищными) почками с мочеточниками, соответствующими вольфовым каналам. В отличие от хрящевых костистые рыбы имеют мочевой пузырь. Что же касается половых протоков костистых рыб, то они представляют собой особые образования, не гомологичные ни вольфовым, ни мюллеровым каналам. Эти особенности возникают в результате изменений хода эмбрионального развития гонад и, по-видимому, связаны с приспособлением к выведению большого количества половых продуктов; плодовитость костистых рыб значительно выше, чем хрящевых. Впрочем, рассмотренные особенности мочеполовой системы являются специфическим свойством только костистых (и некоторых других костных) рыб и не получили дальнейшего развития в эволюции позвоночных животных.
Перечисленные морфологические особенности, а также и ряд прогрессивных физиологических свойств (большая, чем у хрящевых рыб, кислородная емкость крови, некоторые особенности пищеварения, осморегуляции и т. п.) обеспечили костным рыбам возможность значительно более широкой адаптивной радиации как по характеру местообитаний, так и по пищевой специализации.
Костные рыбы заселяют в гидросфере практически все жизненные ниши. Среди этих животных имеются как пелагические морские виды, так и придонные формы, в том числе обитатели больших глубин, хорошо приспособившиеся к высокому давлению и отсутствию освещения. Столь же широко они заселяют и пресные водоемы — как текучие (реки, ручьи), так и стоячие (в том числе и пересыхающие).
Широкий диапазон осморегуляционных процессов позволяет костным рыбам заселять водоемы с разной соленостью и даже в течение индивидуальной жизни переходить из морей в пресные водоемы и наоборот (так называемые проходные рыбы, например многие лососевые). Осморегуляция у костистых рыб построена на совершенно ином принципе, чем у хрящевых. Пресноводные формы имеют хорошо развитую систему фильтрации воды в почках и выделяют большое количество жидкой мочи, избавляя тем самым организм от избытка воды, постоянно поступающей осмотическим путем через жабры и кожу. Морские рыбы, организм которых в силу разницы его осмотического давления с таковым среды все время обезвоживается, имеют слабо развитую клубочковую систему в почках, выделяют небольшое количество сильно концентрированной мочи и пьют много морской воды, выделяя избыток получаемых при этом солей через почки и через жабры.
Ряд физиологических адаптации позволяет костным рыбам жить в водоемах, резко отличающихся содержанием кислорода, температурой и другими условиями.
Очень многообразны адаптации костных рыб к различным типам и способам питания. Этот класс включает и плотоядные, и растительноядные формы (среди хрящевых рыб — только плотоядные); многие виды характеризуются смешанным питанием. Среди костных рыб имеются хищники (которые активно охотятся за крупной добычей или подстерегают ее в засаде) и питающиеся микроскопическими планктонными организмами. Разные виды рыб добывают пищу на поверхности воды, в толще ее, среди подводных зарослей, камней, в коралловых рифах, на поверхности дна и в толще ила и т. д. Все эти формы обладают специальными приспособлениями в строении вторичных челюстей, форме тела, устройстве плавников, окраске и т. п.
Комплекс прогрессивных особенностей строения костных рыб особенно отчетливо и полно выражен у наиболее молодой и прогрессивной ветви этого класса — костистых рыб Teleostei, которая включает подавляющее большинство ныне живущих форм этого класса (около 20 000 видов).
Дополнительная литература
Гуртовой Н. Н., Матвеев Б. С., Дзержинский Ф. Я. Практическая зоотомия позвоночных. Низшие хордовые, бесчелюстные, рыбы. М., 1976.
Никольский Г. В. Частная ихтиология. М., 1971.
Никольский Г. В. Экология рыб. М., 1974.
Строганов Н. С. Экологическая физиология рыб. М., 1962.
Суворов Е. К. Основы ихтиологии. М., 1948.
Шмальгаузен И. И. Основы сравнительной анатомии позвоночных. М., 1947.
Оглавление
ОПИСАНИЕ СКЕЛЕТА
МАТЕРИАЛ И ОБОРУДОВАНИЕ
Тема 7. СКЕЛЕТ КОСТИСТЫХ РЫБ
Т а б л и ц ы: схема расположения костей в черепе костной рыбы; плечевой пояс и скелет грудного плавника судака; тазовый пояс и скелет брюшных плавников судака; скелет хвостового плавника судака.
На одного-двух студентов необходимы:
1. Смонтированный на картонных планшетах разборный скелет
судака.
Смонтированный скелет судака на подставке – один на всю группу.
ЗАДАНИЕ
Рассмотреть скелет в целом. Найти его основные части: череп, позвоночник, скелет парных конечностей и их поясов, скелет непарных конечностей. Рассмотреть детали строения отдельных частей скелета. Сделать следующие рисунки:
2. схема расположения костей в черепе костной рыбы.
3. Грудной пояс с плавником.
4. Тазовый пояс с плавником.
5. Хвостовой плавник.
Осевой скелет (позвоночник). Осевой скелет костистых рыб составлен многочисленными костными позвонками. Тела позвонков спереди и сзади вогнутые – такие позвонки называют амфицельными. Пространство, образующееся между вогнутыми поверхностями соседних позвонков, и узкий канал, пронизывающий в центре тела позвонков, заполнены остатками хорды, имеющей четковидную форму. Позвоночник делится на два отдела: туловищный и хвостовой; позвонки этих отделов отличаются своим строением.
Туловищные позвонки (рис. 29, А) имеют округлое в поперечном сечении тело (29, 1), от которого в стороны отходят поперечные отростки (рис. 29, 2). К этим отросткам причленяются ребра (рис. 29, 3), а к некоторым ребрам – тоненькие мышечные косточки (рис. 29, 8).От верхней (спинной) части позвонка отходят верхние дуги (рис. 29, 4),замыкающиеся остистым отростком (рис. 29, 5). В канале, образованном верхними дугами позвонков, располагается спинной мозг.
| Рис. 29. Позвонки судака. А – туловищный позвонок; Б – хвостовой позвонок: 1 – тело позвонка, 2 – поперечный отросток, 3 – ребро, 4 – верхняя дуга, 5 – верхний остистый отросток, 6 – нижняя дуга, 7 – нижний остистый отросток, 8 – мышечная косточка |
Хвостовые позвонки (рис. 29, Б) также имеют тело и верхние дуги, но поперечные отростки их смещены вниз и образуют нижние дуги (рис. 29, 6), замыкающиеся непарным нижним остистым отростком (рис. 29, 7). Канал, образуемый нижними дугами хвостовых позвонков, носит название гемального; в нем располагаются хвостовые артерия и вена. Расположение этих крупных кровеносных сосудов внутри костного канала предохраняет их от сдавливания при сокращениях мощной мускулатуры хвоста.
Череп.Как и у хрящевых рыб, череп костистых рыб состоит из двух отделов: осевого черепа, или мозговой коробки, и лицевого, или висцерального, черепа. Но в отличие от хрящевых череп костистых рыб почти целиком образован костной тканью и состоит из многочисленных отдельных костей.
Мозговой отдел черепа костистых рыб подразделяется на те же отделы, что и у хрящевых рыб. В каждом отделе образуется несколько костей. Затылочный отдел черепа составлен четырьмя костями: основной, или нижней, затылочной (рис. 30, 3), двумя боковыми затылочными (рис. 30, 10) и верхней затылочной (рис. 30, 26). Эти кости окаймляют большое затылочное отверстие, через которое головной мозг соединяется со спинным. В слуховом отделе черепа располагается по пять ушных костей (рис. 30, 29–33) с каждой стороны. В области глазницы имеются парные боковые клиновидные (рис. 30, 15) и глазоклиновидные кости (рис. 30, 20). Последние расположены на тонкой межглазничной перегородке и при препарировке, как правило разрушаются; у судака их нет. В основании мозговой коробки образуется основная клиновидная кость (рис. 30, 4); при рассматривании черепа сбоку виден лишь ее передний конец.Впереди, в области обонятельного отдела, находятся парные боковые обонятельные (рис. 30, 7) и непарная средняя обонятельная (рис. 30, 16) кости. Все эти кости по происхождению замещающие (первичные), они возникли путем замещения соответствующих участков хрящевого черепа костной тканью.
Рис. 30. Схема расположения костей в черепе костистой рыбы. Основные кости и хрящ заштрихованы, покровные кости – белые: 1 – угловая, 2 – сочленовная, 3 – основная затылочная, 4 – основная клиновидная, ушные кости, 5 – копула, 6 – зубная, 7 – боковая обонятельная, 8 – наружная крыловидная, 9 –внутренняякрыловидная, 10 – боковая затылочная, 11 – лобная, 12 – подвесок, 13 – гиоид, 14 – окостеневшая связка, 15 – боковая клиновидная, 16 – средняя обонятельная, 17 – задняя крыловидная, 18 – верхнечелюстная, 19 – носовая, 20 – глазоклиновидная, 21 – теменная, 22 – небная, 23 – предчелюстная, 24 – парасфеноид, 26 – верхняя затылочная, 27 – дополнительная – symplecti- cum, 28 – сошник, 29–33 – ушные кости, I–V – жаберные дуги
Кости, прикрывающие череп сверху и снизу, по происхождению относятся к покровным (вторичным) костям. Они закладываются в соединительнотканном слое кожи и позже, погружаясь под кожу, прирастают к хрящевой основе черепа. Среди них в крыше черепа заметны крупные плоские лобные кости (рис. 30, 11), покрывающие большую часть черепной коробки. Сзади от них по обе стороны от гребня верхней затылочной располагаются относительно небольшие теменные кости (рис. 30, 21).Спереди от лобных костей в области обонятельного отдела находятся парные носовые кости (рис. 30, 19), разделенные уже упоминавшейся средней обонятельной костью.
Дно черепа прикрыто крупной непарной костью, называемой парасфеноидом (рис. 30, 24), впереди которого расположен непарный сошник (рис. 30, 28), в своей передней части снабженный зубами.
При рассматривании черепа сбоку помимо уже отмеченных костей видно полукольцо из мелких косточек, окаймляющих глазницу – окологлазничных (рис. 31, 1). Передняя из них носит особое название – слезная кость (рис. 31, 2). Все эти кости покровные.
Висцеральный череп костистых рыб представлен серией висцеральных дуг: челюстной (самая передняя), подъязычной и пятью жаберными, из которых последняя (задняя) заметно редуцирована.
Рис. 31. Череп судака сбоку: 1 – окологлазничные кости, 2 – слезная кость, 3 – небная кость, 4 – квадратная кость, 5 – крыловидные кости. 6 – сочленовная кость, 7 – предчелюстная кость, 8 – верхнечелюстная кость, 9 – зубная кость, 10 – угловая кость, 11 – подвесок, 12 – симплектикум (дополнительная), 13 – гиоид, 14 – копула подъязычной дуги, 15 – крышечная кость, 16 – предкрышечная кость, 17 – подкрышечная кость, 18 – межкрышеч- ная кость
В челюстной дуге можно различить первичные и вторичные челюсти. Первичные челюсти образованы преимущественно замещающими костями и гомологичны хрящам небно-квадратному и меккелеву хрящевых рыб. В верхней челюсти они представлены спереди смешанной небной костью (рис. 30, 22, рис. 31, 3), сзади (в месте сочленения с нижней челюстью) – квадратной костью (рис. 30, 25, рис. 31, 4); между ними расположены три крыловидные кости (рис. 29, 5). Из них лишь одна – задняя крыловидная (рис. 30, 17) замещающая, тогда как остальные две – наружная крыловидная (рис. 30, 8) и внутренняя крыловидная (рис. 30, 9,) – по происхождению покровные.
В нижней челюсти первична лишь одна сочленовная кость (рис. 30, 2, рис. 31, 6), осуществляющая подвижное причленение нижней челюсти к верхней (через квадратную кость).
Вторичные челюсти представлены предчелюстными (рис. 30, 23, рис. 31, 7) и верхнечелюстными (рис. 30, 18, рис. 31, 8) костями в верхней половине и крупной зубной костью (рис. 30, 6, рис. 31, 9) – в нижней. Кроме того, в нижней челюсти имеется еще маленькая угловая кость (рис. 30, 1, рис. 31, 10), приросшая к задне-нижнему краю сочленовной. Предчелюстная и зубная кости снабжены довольно крупными зубами. Вторичным челюстям принадлежит главная хватательная функция. Дополнительную функцию удерживания добычи выполняют зубы, расположенные на нёбных костях и сошнике.
Все перечисленные кости челюстной дуги – парные; они симметрично повторяются на правой и левой половинах челюстей.
Подъязычная дуга целиком составлена первичными (замещающими) костями. Верхний элемент ее – подвесок (рис. 30, 12, рис. 31, 11) верхним концом причленяется к слуховому отделу осевого черепа, а нижним – через маленькую дополнительную косточку – symplecticum (рис. 30, 27, рис. 31, 12) – к квадратной кости челюстной дуги. Кроме этого, подвесок срастается с задней крыловидной костью. Таким образом, подвесок выполняет функцию причленения челюстной дуги к осевому черепу; иными словами, череп у костистых рыб, как и у акуловых, гиостилический. Нижняя ветвь подъязычной дуги, гомологичная гиоидному хрящу акуловых рыб, представлена несколькими окостенениями. Из них крупный – гиоид (рис. 30, 13, рис. 31, 13). Гиоиды правой и левой сторон соединяются через непарную кость (рис. 30, 5, рис. 31, 14), одновременно поддерживающую язык. К гиоиду прикрепляются тонкие изогнутые косточки – лучи жаберной перепонки, поддерживающие кожистый край жаберной крышки.
Жаберные дуги представлены каждая четырьмя парными косточками, подвижно сочлененными между собой и объединяющимися снизу при помощи непарных костных элементов – копул. Исключение составляет лишь пятая, самая задняя жаберная дуга, имеющая лишь один парный (нижний) отдел.
Жаберные крышки являются по сравнению с хрящевыми рыбами новообразованием. Каждая состоит из четырех вторичных (покровных) костей: крышки (рис. 31, 15), предкрышки (рис. 31, 16), подкрышки (рис. 31, 17) и межкрышки (рис. 31, 18). Жаберная крышка каждой стороны через предкрышечную кость прикрепляется к соответствующему подвеску и к квадратной кости.
Парные конечности и их пояса.Парные конечности представлены грудными и брюшными плавниками. Опорой грудных плавников в теле рыбы служит плечевой пояс (рис. 33). Он представлен двумя небольшими замещающими (первичными) и несколькими покровными костями. Верхняя из замещающих костей – лопатка (рис. 33, 1) – расположена в области причленения свободной конечности (ее легко отличить по небольшому круглому отверстию в центре кости). Сразу под ней находится коракоид (рис. 33, 2). Этидва элемента составляют первичный пояс. Они неподвижно соединены с крупной покровной костью клейтрум (рис. 33, 5), верхний конец которой направлен несколько вперед; к нему присоединяется небольшая кость надклейтрум (рис. 33, 4).
Клейтрум в свою очередь соединяется с заднетеменной костью (рис. 33, 5). Направленные вперед нижние концы правого и левого клейтрумов соединяются друг с другом. Позади клейтрума неподалеку от лопатки и коракоида расположена небольшая заднеключичная кость (рис. 33, 6). Все названные кости парные; они составляют вторичный плечевой пояс. Правая и левая заднетеменные кости причленяются к осевому черепу, что обеспечивает более прочную фиксацию пояса и таким образом усиливает его опорную функцию.
| Рис. 33. Плечевой пояс и грудной плавник судака: 1 – лопатка, 2 – коракоид, 3 – клейтрум, 4 – надклейтрум, 5 – заднетеменная кость, 6 – заднеключичная кость, 7 – ра- диалии, 8 – костные кожные лучи | Рис. 34. Тазовый пояс и брюшной плавник судака: 1 – тазовые кости, 2 – кожные костные лучи |
Грудной плавник в своем основании имеет один ряд мелких косточек – радиалий (рис. 33, 7), отходящих от лопатки (частично и от коракоида). Вся свободная лопасть плавника состоит из членистых кожных лучей (рис. 33, 8) (эти лучи составлены из мелких чешуеобразных косточек кожного происхождения; отсюда и их название) Особенность скелета грудных плавников костистых рыб, по сравнению с хрящевыми, заключается в редукции базалий. Подвижность грудных плавников увеличивается потому, что мышцы прикрепляются к расширенным основаниям кожных лучей, подвижно сочленяющихся с радиалями.
Тазовый пояс (рис. 34) представлен сливающимися друг с другом парными плоскими треугольными костями, лежащими в толще брюшной мускулатуры и не связанными с осевым скелетом. К боковым сторонам тазового пояса причленяются брюшные плавники. У большинства костистых рыб в скелете брюшных плавников отсутствуют базалий и полностью редуцировались радиалии: лопасть плавника поддерживается кожными костными лучами, расширенные основания которых непосредственно причленяются к тазовому поясу. Такое упрощение скелета брюшных плавников, видимо, связано с их ограниченными функциями.
Непарные конечности. Непарные конечности представлены спинными (у судака их два; некоторые виды имеют только один или три и даже больше спинных плавников), подхвостовым (анальным) и хвостовым плавниками. Анальный и спинные плавники состоят из костных лучей, подразделяющихся на внутренние (скрытые в толще мускулатуры) птеригофоры и наружные плавниковые лучи – лепидотрихии.
| Рис. 35. Хвостовой плавник судака: 1 – конец позвоночного cтолба – уростиль, 2 – гипуралии, 3 – костные кожные лучи – лепидотрихии |
Хвостовой плавник, как это было видно уже при наружном осмотре, имеет внешне равнолопастное строение, однако при рассмотрении его внутреннего скелета (рис. 35) видно, что концевые позвонки позвоночного столба сливаются в палочковидную косточку – уростиль (рис. 35, 1), которая заходит в основание лишь верхней лопасти плавника, а основание нижней лопасти поддерживается разросшимися, довольно широкими нижними дугами позвонков – гипуралиями (рис. 35, 2). Такой тип строения хвостового плавника, вполне или почти симметричного по внешней форме, но асимметричного по расположению внутреннего скелета, носит название гомоцеркального. Наружный скелет хвостового плавника составлен многочисленными кожными лучами – лепидотрихиями (рис. 35, 3).
Строение тела
Вы можете отредактировать эту статью, добавив ссылки на авторитетные источники.
Эта отметка установлена 6 декабря 2017 года.
Покровы
Обладают чешуёй ганоидного, циклоидного или ктеноидного типа. Чешуя покрыта слизью.
Опорно-двигательная система
Скелет костный.
Пищеварительная система
Из ротовой полости пища переходит в глотку, из неё в пищевод, а затем в объёмистый желудок или сразу в кишечник (карась). В желудке происходит частичное переваривание пищи под воздействием желудочного сока. Окончательное переваривание пищи происходит в тонком кишечнике. В начальный отдел тонкого кишечника впадает проток жёлчного пузыря, печени и протоки поджелудочной железы. В тонком кишечнике питательные вещества всасываются в кровь, а непереваренные остатки пищи удаляются через анальное отверстие.
Плавательный пузырь
У большинства костных рыб под позвоночником есть плавательный пузырь, заполненный смесью газов, которые выделяются из кровеносных сосудов. При увеличении объёма пузыря средняя плотность тела рыбы уменьшается и она легко всплывает, а при уменьшении — погружается.
Дыхательная система
Дыхание жаберное. Из ротовой полости вода проходит через жаберные щели, омывает жабры и из-под жаберных крышек выходит наружу. Жабры состоят из жаберных дуг, которые, в свою очередь, состоят из жаберных лепестков и жаберных тычинок.
У некоторых видов существенное значение имеет кожное дыхание или есть приспособления к дыханию воздухом.
Кровеносная система
Кровеносная система рыб замкнутая, сердце состоит из двух камер: предсердия и желудочка. От желудочка к жабрам отходит большой кровеносный сосуд — аорта, ветвящийся на более мелкие — артерии. В жабрах артерии образуют густую сеть мельчайших сосудов — капилляров. После обогащения крови кислородом (обогащённая кислородом кровь называется артериальной) сосуды вновь собираются в артерию, которая ветвится на более мелкие артерии и капилляры. В органах тела через стенки капилляров кислород и питательные вещества поступают в ткани, а из тканей в кровь — углекислый газ и другие продукты жизнедеятельности.
Вы можете отредактировать эту статью, добавив ссылки на авторитетные источники.
Эта отметка установлена 6 декабря 2017 года.
Выделительная система
Ненужные для организма вещества выделяются из крови, когда они проходят через органы выделения — почки. От почек отходят два мочеточника, по которым моча стекает в мочевой пузырь, а затем удаляется наружу через отверстие позади анального.
Органы размножения
У большинства видов оплодотворение наружное. У видов с внутренним оплодотворением копулятивный орган самцов образуется изменённой частью анального плавника. У самцов в парных половых железах, семенниках, образуются сперматозоиды, их называют молоками. У самок в яичниках образуются яйцеклетки, их называют икрой. Оплодотворение, как правило, внешнее, происходит в воде, куда половые клетки выводятся через специальные проливы. Все процессы, связанные с откладыванием икры, молок и оплодотворением, называют нерестом. Для нереста характерно особое брачное поведение.
Классификация
В зоологической литературе XX века группа костных рыб обычно рассматривалась как класс, состоящий из двух подклассов: Лучепёрые (Actinopterygii) и Лопастепёрые (Sarcopterygii). Такая трактовка сохраняется и в некоторых учебниках начала XXI века, однако всё большее распространение получают классификации, в которых ранг упомянутых подклассов повышен до ранга класса. Например, в классификации канадского зоолога Дж. С. Нельсона (2006) группа костных рыб (выступающая под названием Euteleostomi и не имеющая ранга) трактуется как объединение двух сестринских групп — классов Лучепёрые и Лопастепёрые. При этом если лучепёрые являются монофилетической группой, то лопастепёрые — группа заведомо парафилетическая (поскольку ближайший общий предок последней группы также является и предком современных четвероногих); в результате парафилетической оказывается и вся группа костных рыб. Следуя правилам кладистики, не допускающим использования парафилетических таксонов, Нельсон переопределяет объём лопастепёрых, включая в него и всех четвероногих (получивших в его классификации ранг инфракласса).
Примечания
- Наумов С. П. Зоология позвоночных. — М.: Просвещение, 1982. — С. 61—111.
- 1 2 Min Zhu, Wenjin Zhao, Liantao Jia, Jing Lu, Tuo Qiao, Qingming Qu. The oldest articulated osteichthyan reveals mosaic gnathostome characters (англ.) // Nature. — 2009. — Vol. 458, no. 7237. — P. 469-474. — DOI:10.1038/nature07855. — Bibcode: 2009Natur.458..469Z. — PMID 19325627.
- Tyler, David. Critical transitions in fish evolution lack fossil documentation (англ.) // Science Literature : журнал. — 2009. — P. 469—474. Архивировано 12 мая 2013 года.
- Древнейшая костная рыба из Китая. Аммонит.ру. Дата обращения 26 января 2014.
- Michael J. Ryan, Ph.D. Guiyu: The Oldest Articulated Osteichthyan (англ.). PALAEOBLOG. Дата обращения 23 ноября 2009. Архивировано 16 августа 2013 года.
- Descubrimiento de fósil de pez óseo en China aporta nuevos conocimientos clave sobre origen de los vertebrados (исп.). Spanish.china.org.cn. Дата обращения 27 марта 2009. Архивировано 16 августа 2013 года.
- PZ Myers. Guiyu oneiros (англ.). ScienceBlogs. Дата обращения 1 апреля 2009. Архивировано 16 августа 2013 года.
- Helfman, G. S., Facey, D. E. The Diversity of Fishes (неопр.). — Blackwell Sciences, 1997.
- Romer, Alfred Sherwood. The Vertebrate Body. — Philadelphia, PA : Holt-Saunders International, 1977. — P. 396–399. — ISBN 0-03-910284-X.
- Наумов Н. П., Карташёв Н. Н. Зоология позвоночных. Ч. 1. Низшие хордовые, бесчелюстные, рыбы, земноводные. — М.: Высшая школа, 1979. — 333 с. — С. 132.
- Дзержинский Ф. Я., Васильев Б. Д., Малахов В. В. Зоология позвоночных. 2-е изд. — М.: Издат. центр «Академия», 2014. — 464 с. — ISBN 978-5-4468-0459-7. — С. 59.
- Nelson J. S. Fishes of the World. 4th ed. — New York: John Wiley & Sons, 2006. — xx + 601 p. — ISBN 978-0-471-25031-9. — P. 87—88, 461.