Содержание
Лекция № 9. ВЫДЕЛИТЕЛЬНАЯ СИСТЕМА И ВОДНО-СОЛЕВОЙ; ОБМЕН
Выделительная система служит для выведения продуктов обмена и обеспечения водно-солевого состава организма. Основными органами выделения у рыб являются парные туловищные почки с их выводными протоками — мочеточниками, через которые моча поступает в мочевой пузырь. В некоторой степени в экскреции принимают участие кожа, жабры и кишечник.
Почки представляют собой систему выделительных канальцев, открывающихся в общий выводной проток.
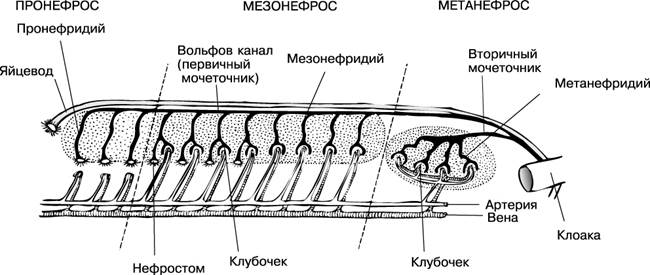
Эволюция выделительной системы у позвоночных происходит в последовательной смене трех типов почек: предпочки, туловищной и тазовой.
Предпочка, или головная почка (пронефрос), развивается в эмбриогенезе у низших позвоночных животных (круглоротые, рыбы, земноводные). Во взрослом состоянии у них функционирует первичная, или туловищная, почка (мезонефрос). У высших позвоночных (амниот) в эмбриогенезе функционирует туловищная почка, а у взрослых — вторичная, или тазовая (метанефрос). Прогрессивное развитие почек связано с усложнением строения почечных канальцев и редукцией мерцательных воронок.
Самой примитивной является головная почка, которая закладывается в передней части полости тела в виде 6-7 выделительных канальцев. Основным фильтрационным элементом ее является воронка, которая одним концом открывается в полость тела, а другим концом — в выводной проток — мочеточник. К верхней части воронки примыкает сосудистый клубочек. Такая почка функционирует у круглоротых и рыб на ранних стадиях развития. У некоторых рыб она сохраняется в виде 2-3 канальцев и во взрослом состоянии (бычки, атерина, бельдюга), а у большинства рыб преобразуется в лимфоидный орган, выполняющий функции кроветворения.
У взрослых круглоротых и рыб позади предпочки развиваются туловищные почки, лентовидные тяжи темно-красного цвета, заполняющие пространство между позвоночником и плавательным пузырем.
Основной функциональной единицей туловищной почки является нефрон, состоящий из мальпигиева тельца и выделительных канальцев. Мальпигиево тельце у рыб малоразмерное (диаметром 50-70 мкм), образовано капсулой Шумлянского — Боумена и сосудистым клубочком. Мочевые канальцы, отходящие от капсул, извиваются, а затем открываются в собирательные канальцы, которые объединяются в более крупные, впадающие затем в мочеточники (рис. 1).
У большинства рыб воронок в почках уже нет, они сохраняются лишь у некоторых хрящевых, а также у осетровых.
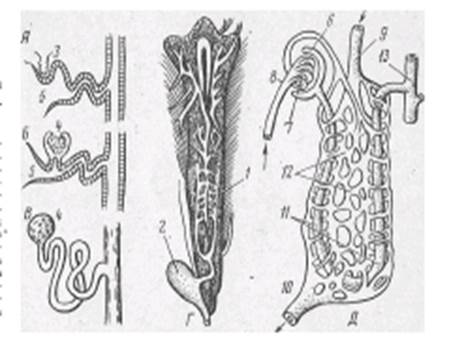 |
Рис.1 Почка форели и схема почечных канальцев рыб: А — головная почка (пред- почка); Б, В, Д— туловищная почка; Г — почка форели; 1 — мочеточники; 2 — мочевой пузырь; 3 — наружный клубочек; 4 — мальпигиево тельце; 5 — воронка; 6 — капсула; 7 — сосудистый клубочек; 8 — ветвь спинной аорты; 9 — воротная вена почек; 10 — почечная вена; 11 — почечные канальцы; 12 — венозные синусоиды; 13 — собирательная трубка. |
 В почках выделяют три отдела: передний, средний и задний, которые у разных рыб имеют разную форму. У всех рыб передний отдел — головная почка. У сазана (карпа) в этом отделе правая и левая почки лежат отдельно, а в заднем срастаются в непарную ленту. Наиболее развит у них средний отдел, сильно расширенный и в виде больших лопастей охватывающий плавательный пузырь. У щуки и окуня передние и задние части почек слиты, а средние лежат раздельно.
В почках выделяют три отдела: передний, средний и задний, которые у разных рыб имеют разную форму. У всех рыб передний отдел — головная почка. У сазана (карпа) в этом отделе правая и левая почки лежат отдельно, а в заднем срастаются в непарную ленту. Наиболее развит у них средний отдел, сильно расширенный и в виде больших лопастей охватывающий плавательный пузырь. У щуки и окуня передние и задние части почек слиты, а средние лежат раздельно.
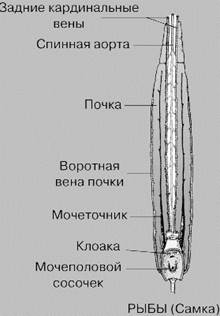
Основным компонентом мочи хрящевых рыб является мочевина, костистых — аммиак, причем аммиак токсичнее мочевины. Почки выполняют в основном роль фильтратора. Продукты обмена веществ доставляются в почки с кровью. От спинной аорты артериальная кровь по почечным артериям поступает в сосудистые клубочки, где происходит ее фильтрация и образуется первичная моча. Выходящие из сосудистых клубочков кровеносные сосуды вместе с сосудами воротной системы почек оплетают выделительные канальцы и, собираясь вместе, образуют задние кардинальные вены. В средней части канальцев происходит обратное всасывание веществ, нужных организму (воды, глюкозы, аминокислот), и образуется вторичная, или окончательная, моча.
Выводным протоком головной почки является пронефрический канал. При развитии туловищной почки пронефрический канал расщепляется на два канала: вольфов и мюллеров. Мюллеров канал у самок хрящевых рыб выполняет функцию яйцевода, у самцов атрофируется. Вольфов канал у круглоротых, костистых и самок хрящевых рыб выполняет функцию мочеточника.

Рис. 2. Мочеполовая система круглоротых и некоторых рыб: I — миноги; II —акулы; III — осетровых; IV — лососевых; V — костистых; 1 — яичник; 2 — яйцевод (мюллеров канал); 3 — вторичный яйцевод; 4 — семенник; 5 — семяпровод; 6 — почка;
7 — мочеточник; 8 — клоака; 9 — мочеполовой синус.
У самцов хрящевых рыб на ранних стадиях развития вольфов канал выполняет одновременно функции мочеточника и семяпровода. У взрослых хрящевых образуется самостоятельный мочеточник, открывающийся в мочеполовой синус, а он в свою очередь — в клоаку, в то время как вольфов канал становится семяпроводом (рис. 2). Для мочеполовой системы костистых в отличие от других рыб характерно отсутствие у них клоаки и полное разделение выделительной и половой систем. Мочеточники (вольфовы каналы) у них по выходе из почек объединяются в непарный проток, который тянется вдоль задней стенки брюшной полости, образуя вырост — мочевой пузырь, заканчивающийся мочевым отверстием, которое открывается позади анального.

Рис. 3. Типы водно-солевого обмена костистых рыб (по Флорикену):
A— пресноводные; Б — морские костистые рыбы.
Строение почек и их функционирование у разных групп рыб связаны с особенностями осморегуляции. У морских хрящевых рыб кровь и тканевые жидкости изотоничны по отношению к окружающей среде, у пресноводных костистых гипертоничны, а у морских костистых гипотоничны. В связи с этим и осморегуляция у них осуществляется по-разному.
У морских хрящевых рыб изотоничность внутренней и внешней среды обеспечивается за счет удержания в крови и тканевых жидкостях мочевины и солей, в результате чего концентрация мочевины в крови у них достигает 2,0—2,5%. Клубочковый аппарат почек развит хорошо, но наружу выводятся лишь излишки мочевины, солей и воды, поэтому количество выделяемой мочи невелико (2—50 мл на 1 кг массы тела в сутки). Для выведения избытка солей у этих рыб есть особая ректальная железа, открывающаяся в прямую кишку.
В связи с тем что у пресноводных рыб осмотическое давление крови и тканевых жидкостей выше, чем в окружающей среде, вода проникает в организм через кожу, жабры, с пищей. Для предупреждения обводнения у них хорошо развит фильтрационный аппарат почек и выделяется большое количество мочи (50—300 мл на 1 кг массы тела в сутки). Потеря солей с мочой компенсируется активной реабсорбцией их в почечных канальцах и поглощением жабрами из воды (рис. 3).
Морские костистые рыбы живут в гипертонической среде, и вода выходит из организма через кожу, жабры, с мочой и фекалиями. Поэтому во избежание иссушения морские костистые рыбы пьют соленую воду, которая из кишечника всасывается в кровь. Часть солей из кишечника удаляется с фекалиями, другая часть выводится секреторными (хлоридными) клетками жаберного аппарата, и таким образом в жидкостях тела поддерживается относительно небольшое осмотическое давление. Клубочковая фильтрация развита слабо, и почки выводят небольшое количество мочи (0,5—20 мл на 1 кг массы тела в сутки). У некоторых рыб в процессе эволюции клубочки исчезают совсем (морская игла, морской черт).
Проходные рыбы при переходе из одной среды в другую могут изменять способ осморегуляции: в морской среде она осуществляется как у морских рыб, а в пресной — как у пресноводных, поддерживая осмотическое давление крови и тканевых жидкостей на определенном уровне.
ПОЛОВАЯ
СИСТЕМА КОСТНЫХ РЫБ
Половая система костных рыб очень своеобразна и совершенно отлична от хрящевых рыб. Мужская половая система состоит из парных вытянутых мешкообразных семенников, от стенок которых внутрь органа отходят многочисленные семенные канальцы, объединяющиеся по направлению к выводному протоку. Если канальцы сильно извиваются в различных плоскостях, то семенник относят к циприноидному типу, такие семенники обычно имеют округлые края, а выводной проток располагается в верхней части органа. Семенники перкоидного типа характеризуются радиальным ходом семенных канальцев, при этом выводной проток находится в центре органа, который на поперечном срезе имеет треугольную форму. Циприноидные семенники имеются у осетровых, карповых, сомовых, щуковых, тресковых, сельдевых и др., а перкоидные — у колюшковых, окуневых и др.
В отличие от хрящевых, семявыносящие протоки семенников половой системы костных рыб самостоятельные, они открываются общим половым отверстием позади анального отверстия. Таким образом, мужские половые протоки костных рыб не имеют никакого отношения к воль- фовым протокам, которые у них служат только мочеточниками.
Женская половая система представлена парными (у самок некоторых видов, например, у окуня, яичник одиночный) удлиненными мешкообразными полыми яичниками, занимающими в теле примерно такое же положение, что и семенники. Часть оболочки яичника вытягивается и образует проток, через который созревшие яйца (их у рыб обычно называют икринками) выходят либо в мочеполовой синус, либо прямо наружу через самостоятельное непарное половое отверстие, лежащее позади анального отверстия. Таким образом, у самок костных рыб мюллеровы протоки редуцируются. У самок разных видов костных рыб могут иметься различные особенности в организации половой системы.
Развитие. У большинства костных рыб оплодотворение внешнее, при этом процесс откладывания яиц (икры) называется нерестом. У некоторых видов он может сопровождаться сложным поведением половых партнеров, особенно сложные ритуалы демонстрируют различные виды цихлид, причем эти рыбы отличаются тем, что выбирают себе партнера (иногда на всю жизнь) в соответствии с собственными взглядами, например, в аквариуме их невозможно принудить вывести потомство с «несимпатичным» представителем противоположного пола. Часто во время нереста изменяется строение рыбы. В отличие от хрящевых, многие виды костных рыб проявляют заботу о потомстве. Это относится к тем же цихлидам, у которых потомство охраняют оба родителя. Самцы лабиринтовых рыб (например, гурами) из пены строят плавучее гнездо, куда помещают икринки (интересно, что самки после икрометания изгоняются самцом и о потомстве не заботятся). Гнездо из травинок строит самец трехиглой колюшки, затем несколько дней присматривает за выклюнувшейся молодью. Интересно заботится о потомстве африканская тиляпия — рыбка вынашивает икринки во рту, но и после выхода молоди в случае опасности мальки быстро заплывают в большой рот родителя и находятся там, пока обстановка не станет спокойной.
Многие виды для нереста мигрируют из моря в реки, обычно туда, где когда-то сами появились на свет (такие рыбы называются проходными). Они демонстрируют удивительные навигационные способности, когда безошибочно находят дорогу за тысячи километров к нужной реке, выбирая ее среди множества других. Полагают, что здесь помогает тонкое обоняние, поскольку лососи сильно возбуждаются, если в окружающую воду добавить воду из родной реки, но остаются невозмутимыми при добавлении воды из реки «чужой». Европейские угри, напротив, во взрослом состоянии живут в реках Европы, но для нереста совершают длительную миграцию в Саргассово море, где на глубине около 200 м нерестятся.
Количество икринок, которое мечет самка, широко варьирует от единичных до астрономических (например, самка луны-рыбы мечет до 30 млн. икринок — рекорд даже среди рыб). Икра некоторых видов свободно плавает в воде, других — прикрепляется к водным растениям, поверхности донных камней и другим подводным предметам, при этом кладка икры у разных видов выглядит неодинаково. Икринки обычно имеют мелкие размеры, покрыты тонкой оболочкой и содержат много желтка, расположенного вблизи одного из полюсов (телолецитальные яйца). Развитие костных рыб чаще всего идет с прохождением личиночной стадии.
Среди костных рыб имеются и живородящие формы, к ним относятся, например, известные аквариумистам гуппи, меченосцы, мол- лиенезии и некоторые другие, при этом количество рожденных детенышей может быть достаточно большим, например, бельдюга рождает 100 — 300 детенышей. Значительная часть икринок поедается другими рыбами, как правило, гибнет много молоди, поэтому до взрослого состояния доживает лишь незначительная часть потомства. При этом замечено, что у менее плодовитых рыб относительная доля выжившей молоди значительно выше, чем у более плодовитых.
Особенности организации костных рыб
Класс Костные рыбы (Osteichthyes)
Общая характеристика костных рыб.
Класс Костные рыбы – самый многочисленный класс позвоночных (более 20 тыс. видов), населяющий все водоемы земного шара.
1) В коже развиваются ганоидные, космоидные или костные чешуи;
2) Внутренний скелет костный или хрящевой (всегда имеются покровные кости);
3) Череп гиостиличный, амфистиличный или аутостиличный;
4) Хвост гомоцеркальный (реже гетероцеркальный или дифицеркальный);
5) Имеется пять пар жаберных щелей, прикрытых общими жаберными крышками;
6) Имеется плавательный пузырь как вырост спинной стороны начальной части пищевода (у некоторых имеются легкие);
7) У большинства вместо артериального конуса в начальной части брюшной аорты образуется луковица аорты;
8) Оплодотворение у большинства наружное;
9) Яйца (икра) не имеют плотной рогоподобной капсулы.
Систематика класса Костные рыбы:
| Подкласс Лопастеперые (Sarcopterygii) | Подкласс Лучеперые (Actinopterygii) | |
| 1. надотряд Кистеперые (Crossopterygomorpha) а) отряд Целакантообразные (Coelacanthiformes) – латимерия | 1. надотряд Ганоидные (Ganoidomorpha) а) отряд Осетрообразные (Acipenseriformes) – белуга, калуга, осетр, стерлядь б) отряд Многоперообразные (Polypteriformes) – многопер в) отряд Амиеобразные (Amiiformes) – ильная рыба г) отряд Панцирникообразные (Lepisosteiformes) – панцирная щука | |
| 2. надотряд Двоякодышащие (Dipneustomorpha) а) отряд Однолегочные (Ceratodiformes) – австралийский рогозуб б) отряд Двулегочные (Lepidosireniformes) – протоптерус, лепидосирен | ||
| 2. группа надотрядов Костистые рыбы (Teleostei) | ||
| 1) надотряд Клюпеоидные (Clupeomorpha) а) отряд Сельдеобразные (Clupeiformes) – сельдевые, анчоусовые б) отряд Лососеобразные (Salmoniformes) – лососевые, хариусовые | ||
| 2) надотряд Ангвилоидные (Anguillomorpha) а) отряд Угреобразные (Anguilliformes) – речные угри, мурены | ||
| 3) надотряд Циприноидные (Cyprinomorpha) а) отряд Карпообразные (Cypriniformes) – харациновидные, гимнотовидные, карповидные б) отряд Сомообразные (Siluriformes) – сом | ||
| 4) надотряд Параперкоидные (Parapercomorpha) а) отряд Трескообразные (Gadiformes) – треска, налим | ||
| 5) надотряд Перкоидные (Percomorpha) а) отряд Окунеобразные (Perciformes) б) отряд Камбалообразные (Pleuronectiformes) |
1) Внешнее строение и покровы:
Кожа представлена многослойным эпидермисом и подстилающим его кориумом. Одноклеточные железы эпидермиса выделяют слизь, имеющую бактерицидное значение и уменьшающую трение. В эпидермисе и кориуме содержатся хроматофорные клетки с пигментами, обуславливающие маскировку (криптическая окраска). Некоторые способны произвольно менять окраску. В кориуме закладываются чешуи костного происхождения:
1. Космоидные чешуи – костные пластинки, покрытые космином (дентиноподобное вещество) (у кистеперых рыб);
2. Ганоидные чешуи – костные пластинки, покрытые ганоином (у ганоидных рыб);
3. Костные чешуи – видоизмененные ганоидные чешуи, у которых исчез ганоин. Типы костных чешуй:
а) Циклоидные чешуи – с гладким краем (карпообразные);
б) Ктеноидные – с зазубренным краем (окунеобразные).
По чешуе можно определить возраст рыбы: в течение года на чешуе образуется два концентрических кольца – широкое, светлое (лето) и узкое, темное (зима). Следовательно, два кольца (полосы) – один год.
2) Внутреннее строение:
а) Пищеварительная система:
— ротовая полость: имеются развитые зубы, нерегулярно сменяющиеся в течение жизни. У некоторых намечается гетеродонтность (неоднородность зубов). Языка нет. Железы выделяют слизь, не содержащую пищевых ферментов, она лишь способствует проталкиванию пищевого комка.
— глотка: в продвижении пищи участвуют жаберные тычинки жаберных дуг. У некоторых они образуют цедильный аппарат (планктоядные), у некоторых способствуют проталкиванию пищи (хищные), или перетирают пищу (бентосоядные).
— пищевод: короткий, мускулистый, незаметно переходит в желудок.
— желудок: разной формы, у некоторых отсутствует. Железы вырабатывают соляную кислоту и пепсин. Следовательно, здесь осуществляется химическая обработка белковой пищи.
— кишечник: спиральный клапан отсутствует. Имеются пилорические выросты в начальной части кишечника, увеличивающие всасывающую и переваривающую поверхность кишечника. Кишечник длиннее, чем у хрящевых рыб (у некоторых в 10-15 раз превышает длину тела). Клоаки нет, кишечник открывается наружу самостоятельным анальным отверстием.
— печень: менее развита (5% от массы тела). Желчный пузырь и проток развит хорошо.
— поджелудочная железа: неоформлена, островками разбросана по стенкам кишечника и печени.
б) Дыхание и газообмен:
Органы дыхания – жабры, состоящие из жаберных лепестков, находятся на 1-4 жаберных дугах (костные). Межжаберные перегородки отсутствуют. Жаберная полость прикрывается костными жаберными крышками. К основанию жаберной дуги подходит приносящая жаберная артерия, дающая капилляры в жаберные лепестки (газообмен); выносящая жаберная артерия собирает из жаберных лепестков окисленную кровь.
Акт дыхания: при вдохе жаберные крышки отходят в стороны, а их кожистые края наружным давление прижаты к жаберной щели и препятствуют выходу воды. Вода насасывается в жаберную полость через ротоглоточную полость и омывает жабры. При выдохе жаберные крышки сближаются, вода давлением открывает края жаберных крышек и выталкивается наружу.
Жабры также участвуют в выделении метаболитов и вводно-солевом обмене.
Кроме жаберного дыхания у некоторых костных рыб развиты:
1. Кожное дыхание (от 10 до 85% в дыхании);
2. С помощью ротовой полости (ее слизистая богата капиллярами);
3. С помощью наджаберного органа (полые камеры над жабрами с развитой складчатостью внутренних стенок);
4. С помощью кишечника (заглатываемый пузырек воздуха проходит по кишечнику, отдавая в кровяное русло О2 и забирая СО2);
5. Плавательный пузырь у открытопузырных рыб (плавательный пузырь связан с пищеводом). Основная роль – гидростатическая, барорецептор и акустический резонатор;
6. Легочное дыхание (у кистеперых и двоякодышащих). Легкие развиваются из плавательного пузыря, стенки которого приобретают ячеистое строение и оплетаются сетью капилляров.
в) Кровеносная система:
Один круг кровообращения, двухкамерное сердце, имеется венозная пазуха. Луковица аорты, замещающая артериальный конус имеет гладкомышечные стенки и, следовательно, к отделам сердца не относится.
Артериальная часть:
Сердце → брюшная аорта → 4 пары приносящих жаберных артерий → капилляры жабр → 4 пары выносящих жаберных артерий → корни спинной аорты → сонный головной круг (к голове) и спинная аорта (к внутренним органам) → хвостовая артерия.
Венозная часть:
Передние кардинальные вены от головы и подключичные вены от грудных плавников → кювьеровы протоки → венозная пазуха → сердце.
Хвостовая вена → воротные вены почек → воротная система почек → задние кардинальные вены → кювьеровы протоки → венозная пазуха → сердце.
От кишечника → воротная вена печени → воротная система печени → печеночная вена → венозная пазуха → сердце.
Кроветворные органы – селезенка и почки.
г) Выделительная система:
Парные мезонефрические почки → мочеточники (вольфовы каналы) → мочевой пузырь → самостоятельное мочевое отверстие.
У пресноводных рыб почки гломерулярные (развиты боуменовы капсулы с мальпигиевыми тельцами). У морских гломерулы уменьшаются и упрощаются. Продукт выделения – аммиак.
2 типа водно-солевого обмена:
а) Пресноводный тип: в связи с гипотоничностью среды вода постоянно поступает в организм через кожу и жабры, следовательно, рыбам грозит обводнение, что приводит к развитию фильтрационного аппарата, позволяющего выводить избыток воды (до 300 мл конечной мочи на 1 кг массы тела в сутки). Потеря солей избегается активной реабсорбцией их в почечных канальцах.
б) Морской тип: в связи с гипертоничностью среды вода выходит из организма через кожу и жабры, следовательно, рыбам грозит обезвоживание, что приводит к развитию агромерулярных почек (исчезают гломерулы) и уменьшению количества конечной мочи до 5 мл на 1 кг массы тела в сутки.
д) Половая система:
♂: Семенники → семявыносящие канальцы → семяпроводы (самостоятельные каналы, не связанные с мезонефросом) → семенной пузырек → половое отверстие.
♀: Яичники → задние вытянутые отделы яичников (выводные протоки) → половое отверстие.
Большинство рыб раздельнополы. Оплодотворение наружное. Самка откладывает икру (яйца), а самец поливает ее молоками (сперматозоиды).
е) Нервная система и органы чувств:
Аналогичны таковым системам хрящевых рыб.
3) Скелет и мышечная система:
Хрящевая ткань замещается костной: образуются основные (замещающие) кости. В кориуме закладываются второй тип костей: покровные (кожные) кости, погружающиеся под кожу и входящие в состав скелета.
а) Осевой скелет:
Представлен хорошо развитыми костными амфицельными позвонками. В телах позвонков и между ними проходит четковидная хорда. Позвоночный столб представлен туловищным и хвостовым отделами, строение которых аналогично хрящевым рыбам. Позвонки соединены с помощью суставных отростков, расположенных у основания верхних дуг.
б) Череп:
1. Мозговой череп.
Характерно наличие большого количества основных и покровных костей.
— в затылочном отделе 4 затылочные кости: основная затылочная, 2 боковые и верхняя затылочная кости.
— боковой отдел образован 5 ушными костями, 3 глазничными костями (глазоклиновидная, основная и боковая клиновидная), 2 обонятельными костями (непарная средняя обонятельная и боковые парные обонятельные). Все эти кости основные: развиваются путем окостенения хряща.
— крыша мозгового черепа образована покровными костями: парными носовыми, лобными и теменными костями.
— дно мозгового черепа образовано 2 непарными кожными костями: парасфеноидом и сошником с зубами.
2. Висцеральный череп:
Челюстная, подъязычная, 5 пар жаберных дуг и скелет жаберной крышки.
— челюстная дуга делится на первичные челюсти – окостенение хрящевых элементов челюстной дуги, и вторичные челюсти – покровные кости, укрепляющие челюсти. Из небно-квадратного хряща (верхняя челюсть) образуются 3 основные кости: небные (с зубами), задние крыловидные и квадратная. Между ними располагаются покровные наружные и внутренние крыловидные кости. Из меккелева хряща (нижняя челюсть) образуется замещающая сочленовная кость, образующая с квадратной костью челюстной сустав. Вторичные челюсти представлены в верхней челюсти предчелюстными и верхнечелюстными костями с зубами; в нижней челюсти – зубной и угловой костями.
— подъязычная дуга образована основными костями: гиомандибуляре, гиоидом и непарной копулой. Для костных рыб характерна гиостилия.
— скелет жаберной крышки представлен 4 покровными костями: предкрышечной, крышечной, межкрышечной и подкрышечной.
— жаберных дуг 5 пар. Первые 4 образованы 4 парными элементами, соединенными снизу копулами (они несут жабры). Последняя жаберная дуга не несет жабр и состоит из 2 парных элементов, к которым могут быть причленены глоточные зубы (у некоторых).
в) Скелет парных конечностей и их поясов:
Парные конечности представлены грудными и брюшными плавниками. Различают 2 типа парных плавников:
а) бисериальный тип – плавники имеют центральную расчлененную ось, к которой попарно прикрепляются членики радиалий (лопастеперые и двоякодышащие);
б) унисериальный тип – радиалии прикрепляются только с одной стороны центральной оси (кистеперые рыбы).
У лучеперых рыб базальные элементы плавников редуцируются, радиалии прикрепляются непосредственно к поясу, к радиалиям прикрепляются лепидотрихии (кожные костные лучи, поддерживающие лопасть плавника).
Плечевой пояс состоит из первичных и вторичных элементов. Первичный пояс представлен окостеневшими лопатками и коракоидом. Вторичный пояс представлен крупным клейтрумом, который посредством супраклейтрума присоединяется к затылочному отделу черепа.
Скелет собственно грудных плавников представлен одним рядом радиалий, к которому крепятся лепидотрихии.
Тазовый пояс представлен хрящевой или костной пластинкой, лежащей в толще мускулатуры, к которой через ряд радиалий крепятся лепидотрихии брюшных плавников.
г) Скелет непарных конечностей:
Спинные плавники образованы лепидотрихиями, скелетной основой которых являются птеригофоры, погруженные в мускулатуру и нижними концами содненные с верхними остистыми отростками позвонков.
Хвостовой плавник: 4 типа:
1. Протоцеркальный – симметричное строение, хорда проходит по середине плавника (личинки рыб).
2. Гетероцеркальный – аналогичен хрящевым рыбам (осетрообразные).
3. Гомоцеркальный – равнолопастной, верхняя и нижняя лопасти одинаковы, но осевой скелет заходит в верхнюю лопасть (большинство костных рыб).
4. Дифицеркальный – однолопастной. Осевой скелет проходит по середине плавника (двоякодышащие и кистеперые рыбы).
Скелетной основой хвостового плавника являются расширенные отростки концевых позвонков – гипуралии, лопасть плавника поддерживают лепидотрихии.
Мышечная система аналогична хрящевым рыбам.
Анисимова И.М., Лавровский В.В.
«Ихтиология»
Из-во Высшая школа. 1983 г.
ГЛАВА I
СТРОЕНИЕ И НЕКОТОРЫЕ ФИЗИОЛОГИЧЕСКИЕ ОСОБЕННОСТИ РЫБ
ПОЛОВАЯ СИСТЕМА
Развитие мочеполовой системы в эволюции рыб привело к обособлению половых протоков от выделительных.
У круглоротых специальных половых протоков нет. Из разрывающейся половой железы половые продукты выпадают в полость тела, из нее – через половые поры – в мочеполовой синус, а затем через мочеполовое отверстие выводятся наружу.
У хрящевых рыб половая система связана с выделительной. У самок большинства видов яйца выводятся из яичников по мюллеровым каналам, выполняющим роль яйцеводов и открывающимся в клоаку; вольфов канал является мочеточником. У самцов вольфов канал служит семяпроводом и через мочеполовой сосочек также открывается в клоаку.
У костистых рыб вольфовы каналы служат мочеточниками, мюллеровы каналы у большинства видов редуцируются, половые продукты выводятся наружу через самостоятельные половые протоки, открывающиеся в мочеполовое или половое отверстие.
У самок (большинства видов) зрелые яйца выводятся из яичника наружу через короткий проток, образованный оболочкой яичника. У самцов канальцы семенника соединяются с семяпроводом (не связанным с почкой), который открывается наружу мочеполовым или половым отверстием.
Половые железы, гонады – семенники у самцов и яичники или ястыки у самок – лентовидные или мешковидные образования, висящие на складках брюшины – брыжейке – в полости тела, над кишечником, под плавательным пузырем. Строение гонад, сходное в основе, у разных групп рыб имеет некоторые особенности, У круглоротых половая железа непарная, у настоящих рыб гонады большей частью парные. Вариации в форме гонад у различных видов главным образом выражаются в частичном или полном слиянии парных желез в одну непарную (самки трески, окуня, бельдюги, самцы песчанки)или в ясно выраженной асимметрии развития: часто гонады бывают разные по объёму и массе (мойва, серебряный карась и др. ), вплоть до полного исчезновения одной из них. С внутренней стороны стенок яичника в щелевидную полость его отходят поперечные яйце несущие пластинки, на которых развиваются половые клетки. Основу пластинок составляют соединительнотканные тяжи с многочисленными ответвлениями. Вдоль тяжей проходят сильно разветвленные кровеносные сосуды. Зрелые половые клетки выпадают с яйценесущих пластинок в полость яичника, которая может быть расположена в центре его (например, окуневые) или сбоку (например, карповые).
Яичник непосредственно сливается с яйцеводом, выводящим яйца наружу. У некоторых форм (лососевые, корюшковые, угревые) яичники не замкнуты и зрелые яйца выпадают в полость тела, а уже из неё через специальные протоки выводятся из организма. Семенники большинства рыб – парные мешковидные образования. Зрелые половые клетки по выводным протокам – семяпроводам – выводятся во внешнюю среду через специальное половое отверстие (у самцов лососей, сельдей, щуки и некоторых других) или через мочеполовое отверстие, расположенное позади ануса (у самцов большинства костистых рыб).
У акул, скатов, химер имеются придаточные половые железы (передняя часть почки, становящаяся лейдиговым органом); выделения железы примешиваются к сперме.
У некоторых рыб конец семяпровода расширен и образует семенной пузырек (не гомологичен органам того же названия у высших позвоночных).
Известно о железистой функции семенного пузырька у некоторых представителей костистых рыб. От внутренних стенок семенника отходят внутрь семенные канальцы, сходящиеся к выводному протоку. По расположению канальцев семенники костистых рыб разделяют на две группы: циприноидные, или ацинозные, – у карповых, сельдевых, лососевых, сомовых, щуковых, осетровых, тресковых и др. ; перкоидные, или радиальные, – у окуневых, колюшковых и др. (рис. 24).
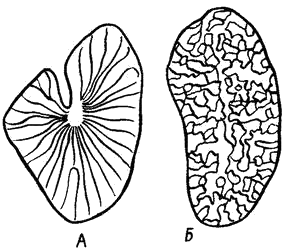
Рис. 24. Типы строения семенников костистых рыб
А – перкоидный; Б – циприноидный
В семенниках циприноидного типа семенные канальцы извиваются в различных плоскостях и без определённой системы. Вследствие этого на поперечных гистологических срезах видны их отдельные участки неправильной формы (так называемые ампулы). Выводной проток помещается в верхней части семенника. Края семенника округлы.
В семенниках перкоидного типа семенные канальцы тянутся от стенок семенника радиально. Они прямые, выводной проток расположен в центре семенника. Семенник на поперечном срезе имеет треугольную форму.
По стенкам канальцев (ампул) лежат крупные клетки – исходные семенные клетки, первичные сперматогонии, будущие сперматозоиды.
Половые клетки появляются на ранних этапах развития у эмбрионов в генитальных складках, которые тянутся вдоль полости тела. У молоди лососей (горбуша, кета, нерка, сима, кижуч и атлантический лосось) первичные половые клетки обнаруживаются на стадии формирования первично-почечных протоков. У зародыша атлантического лосося первичные половые клетки были выявлены в возрасте 26 cyт. У мальков рыб уже можно найти половые железы в виде волосовидных тяжей.
Овогонии – будущие икринки – образуются в результате деления зачатковых клеток зародышевого эпителия, это округлые, очень мелкие, не видимые простым глазом клетки. После овогониальных делений овогония превращается в овоцит. В дальнейшем в ходе овогенеза – развития яйцевых клеток – различают три периода: период синаптенного пути, период роста (малого – протоплазматического и большого – трофоплазматического) и период созревания.
Каждый из этих периодов подразделяется на ряд фаз. Период синаптенного пути характеризуется главным образом преобразованием ядра клетки (овоцита). Затем наступает период малого –протоплазматического – роста, когда увеличение размеров овоцита происходит за счет накопления цитоплазмы. Здесь в развитии овоцитов выделяют ювенильную фазу и фазу однослойного фолликула.
В ювенильной фазе овоциты ещё относительно небольшие, чаще всего округлой формы, с тонкой, бесструктурной, так называемой первичной (вырабатываемой самой яйцеклеткой) оболочкой, к которой прилегают отдельные фолликулярные клетки, а снаружи – клетки соединительной ткани. Ядро овоцита имеет хорошо заметную тонкую оболочку; округлое большое, оно почти всегда лежит в центре. По периферии ядра располагаются многочисленные ядрышки, в большинстве прилегающие к оболочке. В фазе однослойного фолликула собственная оболочка становится толще, над ней формируется фолликулярная оболочка с прилегающими отдельными соединительнотканными клетками.
В этой же фазе в овоците часто можно обнаружить вителлогенную зону. Эта зона имеет ячеистую, как бы пенистую структуру и возникает в цитоплазме вокруг ядра, на некотором расстоянии от него (циркумнуклеарная зона). К концу фазы (и периода) овоциты увеличиваются настолько, что их можно различить с помощью лупы или даже невооруженным глазом.
На протяжении формирования яйцевой клетки наряду с преобразованиями ядра в ней образуются и накапливаются питательные вещества, концентрирующиеся в желтке (белки и липиды) и чисто липидных включениях, которые затем, в период развития эмбриона, используются для его пластических и энергетических нужд. Этот процесс начинается в периоде большого роста овоцита, когда на периферии его появляются вакуоли, содержащие углеводы. Таким образом, период большого (трофоплазматического) роста овоцита характеризуется увеличением не только количества протоплазмы, но накоплением в нем питательных, трофических веществ – белка и жира.
В период большого роста происходят вакуолизация цитоплазмы, появление желтка и наполнение им овоцита. Период большого роста также состоит из нескольких фаз. В фазе вакуолизации цитоплазмы овоциты, увеличенные по сравнению с предыдущей фазой, имеют несколько угловатую форму вследствие давления соседних клеток. Оболочки овоцита – собственная, фолликулярная, соединительнотканная – стали четче выраженными. На периферии овоцита образуются единичные мелкие вакуоли, которые, увеличиваясь в числе, создают более или менее густой слой. Это будущие кортикальные альвеолы, или гранулы. Содержимое вакуолей составляют углеводы (полисахариды), которые после оплодотворения яйца способствуют всасыванию под оболочку воды и образованию перивителлинового пространства. У некоторых видов (лосось, карп) раньше вакуолей в цитоплазме появляются жировые включения. В ядре ядрышки отходят от оболочки вглубь. В следующей фазе – первоначального накопления желтка – на периферии овоцита между вакуолями появляются отдельные мелкие шарики желтка, количество которых быстро растет, так что к концу фазы они занимают почти всю плазму овоцита.
В собственной оболочке появляются тонкие канальцы, придающие ей радиальную исчерченность (Zona radiata); по ним в овоцит проникают питательные вещества. Над собственной оболочкой у некоторых рыб образуется ещё одна вторичная – оболочка (производная фолликулярных клеток, окружающих овоцит). Эта оболочка, разнообразная по структуре (студенистая, сотовая или ворсинчатая), после выхода овоцита из фолликула служит для прикрепления икринки к субстрату. Фолликулярная оболочка становится двуслойной. Границы ядра отчетливы, но стали извилистыми, ‛лапчатыми“.
Следующая фаза – наполнение овоцита желтком – характеризуется очень сильным увеличением объёма желтка, частицы которого приобретают вместо шаровидной многогранную, глыбковидную форму. Вакуоли отжимаются к поверхности овоцита.
Из-за преобладания в это время количественных изменений (без существенных морфологических сдвигов) некоторые исследователи считают нецелесообразным выделять эту фазу как самостоятельную. К концу фазы овоцит достигает дефинитивных размеров. Заметны изменения желтка и ядра: ядро начинает смещаться (к анимальному полюсу), его контуры становятся менее ясными; частицы желтка начинают сливаться. Заканчивается формирование вторичной оболочки.
Последняя фаза развития – фаза зрелого овоцита. Частицы желтка у большинства рыб (за исключением вьюна, макропода, некоторых карповых) сливаются в гомогенную массу, овоцит становится прозрачным, цитоплазма сосредоточивается на периферии овоцита, ядро теряет контуры.
Преобразования ядра вступают в завершающую стадию.
Одно за другим следуют два деления созревания. В результате образуются ядро зрелого овоцита с гаплоидным числом хромосом и три редукционных тельца, которые не участвуют в дальнейшем развитии, отделяются от яйца и дегенерируют. После второго деления созревания митотическое развитие ядра доходит до метафазы и в этом состоянии остается до оплодотворения.
Дальнейшее развитие (формирование женского пронуклеуса и отделение полярного тельца) происходит уже после оплодотворения.
Сквозь собственную (Z. radiata) и студенистую оболочку проходит канал (микропиле), через который сперматозоид проникает в яйцеклетку при оплодотворении. У костистых рыб одно микропиле, у осетровых – несколько: у севрюги – до 13, белуги – до 33, черноморско-азовского осетра – до 52. Поэтому полиспермия возможна только у осетровых рыб, но не у костистых.
При овуляции фолликулярная и соединительнотканная оболочки лопаются и остаются на яйценесущих пластинках, а освобожденный из них овоцит, окруженный собственной и студенистой оболочками, выпадает в полость яичника или полость тела. Здесь овулировавшие яйца находятся в полостной (овариальной) жидкости, сохраняя относительно длительное время способность к оплодотворению (табл. 3). В воде или вне полостной жидкости они эту способность быстро теряют.
У акул и скатов, которым свойственно внутреннее оплодотворение, оплодотворенное яйцо, продвигаясь по половым путям, окружается ещё одной – третичной – оболочкой. Рогоподобное вещество этой оболочки образует твердую капсулу, надежно защищающую зародыш во внешней среде (см. рис. 34).
В процессе развития овоцитов наряду с другими изменениями происходит колоссальное увеличение его размеров: так, по сравнению с овогониями, образованными при последнем овогониальном делении, объём зрелого овоцита возрастает у окуня в 1 049 440, у воблы – в 1 271 400 раз.
У одной самки овоциты (а после овуляции – икринки) неодинаковы по величине: самые крупные могут превосходить самые мелкие в 1,5–2 раза. Это зависит от их расположения на яйценосной пластинке: овоциты, лежащие вблизи кровеносных сосудов, лучше снабжаются питательными веществами и достигают больших размеров.
Отличительной особенностью процесса развития сперматозоидов – сперматогенеза – является многократное уменьшение клеток. Каждый исходный сперматогоний делится несколько раз, в результате чего возникает скопление сперматогониев под одной оболочкой, называемое цистой (стадия размножения). Образовавшийся при последнем делении сперматогоний несколько увеличивается, в его ядре происходят мейотические преобразования и сперматогоний превращается в сперматоцит I порядка (стадия роста). Затем наступают два последовательных деления (стадия созревания): сперматоцит I порядка делится на два сперматоцита II порядка, вследствие деления которых образуются две сперматиды. В следующей – завершающей – стадии формирования сперматиды превращаются в сперматозоиды. Таким образом, из каждого сперматоцита образуются четыре сперматиды с половинным (гаплоидным)набором хромосом. Оболочка цисты лопается, и сперматозоиды наполняют семенной каналец. Через семяпроводы созревшие сперматозоиды выходят из семенника, а затем по протоку – наружу.
Характерными особенностями развития семенников является сильная неравномерность (асинхронность) развития органа в целом. Особенно сильно эта неравномерность проявляется у впервые созревающих рыб, но вполне отчетливо выражена и у нерестующих повторно половозрелых особей. В результате практически все самцы нерестуют порционно и в течение длительного периода от них можно получать сперму.
Процесс созревания половых клеток у разных рыб проходит, в общем, по одной схеме. По мере развития половых клеток внутри яичников и семенников меняются и внешний вид, и размеры гонад. Это побудило составить так называемую шкалу зрелости гонад, пользуясь которой можно было бы по внешним признакам гонад определить степень зрелости половых продуктов, что очень важно в научных и промысловых исследованиях. Чаще других используют универсальную 6-балльную шкалу, в основе которой лежат общие признаки для разных видов рыб (табл. 4, 5; рис. 25, 26).
Предложены также и другие шкалы, в которых учитывают особенности созревания определённых групп рыб. Так, для яичников карповых и окуневых В. М. Мейеном предложена 6-балльная шкала, а для семенников С. И. Кулаевым – 8-балльная.
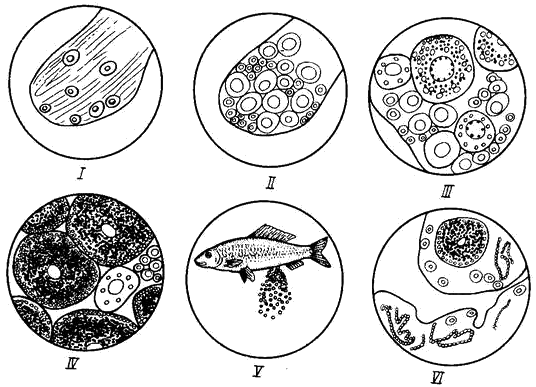
Рис. 25. Стадии (I – VI) зрелости гонад самок костистых рыб 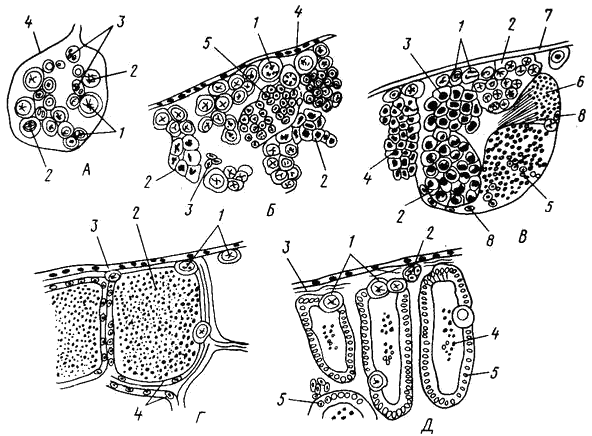
Рис. 26. Стадии зрелости гонад самцов костистых рыб (по Сакун, Буцкой, 1968):
А – I стадия (1 – сперматогоний, 2 – делящийся сперматогоний, 3 – кровеносный сосудс эритроцитами, 4 – оболочка семенника); Б – II стадия (1 — сперматогоний, 2 – делящийся сперматогоний, 3 – кровеносный сосуд, 4 – оболочка, семенника, 5 – циста с мелкими сперматогониями); В – III стадия (1 – сперматогоний, 2 – циста со сперматоцитами I порядка, 3 – циста с делящимися сперматоцитами I порядка, 4 – циста с делящимися сперматоцитами II порядка, 5 – циста со сперматидами, 6 – циста со зрелыми сперматозоидами, 7 – оболочка семенника, 8 – фолликулярный эпителий); Г – IV стадия (1 – сперматогоний, 2 – сперматозоиды, 3 – оболочка семенника, 4 – фолликулярный эпителий); Д – VI стадия (1 – сперматогоний, 2 кровеносный сосуд, 3 – оболочка семенника, 4 – остаточные сперматозоиды, 5 – фолликулярный эпителий)
У подавляющего большинства рыб осеменение наружное. У хрящевых рыб, которым свойственны внутреннее осеменение и живорождение, имеются соответствующие изменения в строении полового аппарата. Развитие зародышей у них происходит в заднем отделе яйцеводов, получивших название матки. Из костистых рыб живорождение свойственно гамбузии, морскому окуню, многим аквариумным рыбкам. У них молодь развивается в яичнике.
 Оглавление
Оглавление