
Содержание
- Лимфатические узлы животных и особенности их строения
- Изучение лимфатической системы и топографии лимфатических узлов у различных видов сельскохозяйственных животных по атласам и методическим рекомендациям.
- Сердечно-сосудистая и лимфатическая системы птиц
- Лимфатическая система
- Лимфатическая система птичьего эмбриона
Лимфатические узлы животных и особенности их строения
 24.10.2015
24.10.2015
Лимфатические узлы, как правило, имеют бобовидную форму располагаются группами (пакетами) по ходу лимфатических сосудов в различных частях тела (регионально) и в зависимости от дренируемой ими области именуются подчелюстными, околоушными, подколенными, средостенными, брыжеечными и т.д.
Форма лимфатических узлов разнообразна, но каждый из них имеет вогнутую и выпуклую поверхность. Через вогнутую поверхность лимфатические сосуды входят в узел, а через выпуклую (ворота органа) — выходят. Только у свиней, наоборот, в выпуклую поверхность (ворота органа) проникают афферентные лимфатические сосуды, а через вогнутую поверхность выходят эфферентные.
Размеры и количество лимфатических узлов различаются в широких пределах. Величина лимфатических узлов колеблется от 1 мм до 10-12 см в длину, под влиянием антигенного стимула размеры узлов увеличиваются до 15 раз. Их количество также сильно варьирует — до 300 у крупного рогатого скота, до 8000 — у лошадей, около 190 — у свиней, около 60 — у собак. У человека их насчитывают от 500 до 1000. Число узлов в группах составляет от 1-3 до 20 и более.
Относительная масса лимфатических узлов животных колеблется в пределах сотых или тысячных долей процента общей массы тела.
Роль лимфатических узлов в иммунном ответе в существенной степени определяется местом аппликации антигена. При подкожном его введении высокую значимость в продукции антител имеют региональные лимфатические узлы. В случае внутривенной иммунизации основная роль в выработке антител принадлежит селезенке.
Функции лимфатических узлов
Находясь на путях тока лимфы, лимфатические узлы выполняют функции своеобразных биологических фильтров, задерживающих разносимые по организму микробы, погибшие и опухолевые клетки, антигены и другие крупные чужеродные частицы. Они попадают в узел с лимфой (тканевой жидкостью), доставляемой приносящими (афферентными) сосудами (обычно 2-4), которые начинаются в тканях капиллярной сетью. В лимфатическом узле чужеродные частицы фрагментируются фагоцитами, распознаются клетками системы иммунитета, индуцируют формирование иммунного ответа. Местами скопления лимфы для ее более тщательной очистки являются синусы — узкие щели, стенки которых выстланы клетками плоского эпителия. Внутренняя часть синусов представлена мелкопетлистой сетью ретикулярных клеток и волокон. Благодаря синусам, под влиянием осмотического давления и сокращения мышц обеспечивается ток лимфы через лимфатический узел. Притекающая в узел лимфа через афферентные сосуды попадает в субкапсулярный синус, расположенный под капсулой узла над его паренхиматозной кортикальной зоной (рис, 1.8). Из субкапсулярного синуса лимфа попадает в паренхиму, сначала в промежуточные синусы коркового слоя, а затем в синусы мозговой области и после этого в воротные синусы, из которых выходят эфферентные лимфатические сосуды на противоположной от афферентных сосудов стороне, в области ворот узла (небольшое вдавление капсулы). Через ворота выходят также вены, входят нервы и артерии. Оттекающая из узла лимфа собирается в крупные лимфатические сосуды и через грудной проток попадает в верхнюю полую вену. Лимфатические капилляры, сосуды, проток формируют лимфатическую систему, через которую, как отмечалось выше, профильтровывается тканевая жидкость из всех регионов тела и возвращается в кровеносное русло.

Помимо фильтрационной лимфатическая система выполняет транспортные функции, благодаря которой малые лимфоциты (основные и фактически единственные клетки лимфы) попадают в кровеносное русло, с током крови разносятся по организму, через систему капилляров и сосудов возвращаются в лимфатические узлы и снова попадают через грудной проток в кровеносное русло. Считается, что не менее 95% лимфоцитов лимфатического узла составляют клетки, пришедшие в узел благодаря циркуляторным процессам. В подколенном лимфатическом узле овцы, например, популяция лимфоцитов полностью обновляется за 70 час. Процессы рециркуляции лимфоцитов имеют важнейшее значение для системы иммунитета, которая благодаря этим процессам функционирует как единое целое.
Строение лимфатического узла
Схематическое строение лимфатического узла показано на рис. 1.8. С наружной стороны лимфатический узел покрыт соединительнотканной капсулой, от которой внутрь органа отходят тонкие соединительнотканные перегородки (трабекулы). В области ворот (хилус) от капсулы в паренхиму узла отходят короткие более толстые соединительнотканные ответвления, которые могут соединяться с трабекулами и формировать дольчатое строение лимфатического узла.
Обычно лимфатические узлы имеют одни ворота, однако некоторые из них, располагающиеся, например, на путях тока лимфы от внутренних органов, могут иметь до 3-4 ворот. Другие узлы, например брыжеечные, характеризуются большим относительным содержанием В-лимфоцитов, в частности, с мембранным IgA. Однако в целом по строению и функциям лимфатические узлы разной локализации мало отличаются друг от друга.
Паренхима лимфатического узла представлена ретикулярной тканью (специализированный вид соединительной ткани, состоящей из клеток, соединенных между собой отростками, и межклеточного вещества — ретикулярные волокна, тесно соединенные с клетками), в петлях трехмерной сети которой локализуются клеточные элементы, преимущественно лимфоциты на разных стадиях развития и функционирования. Паренхиму подразделяют на наружную корковую и более глубокую, мозговую области. Между корковой и мозговой областями локализуется паракортикальная зона (рис. 1.8а).
Кортикальная зона узла содержит диффузную лимфоидную ткань и лимфоидные узелки — фолликулы, в которых сосредоточены В-лимфоциты.
Вследствие этого эту зону именуют В-зависимой, а фолликулы — первичными фолликулами. В условиях формирования иммунного ответа в первичных фолликулах появляются центры размножения, или зародышевые центры, включающие пролиферирующие В-лимфобласты и густую сеть специализированных фолликулярных дендритных клеток, создающих структурную основу фолликулов. Фолликул, содержащий зародышевые центры, называют вторичным фолликулом. Периферическая область вторичного фолликула (мантия), окружающая зародышевый центр, состоит из концентрического слоя плотно упакованных В-лимфоцитов с мембранными иммуноглобуинами — IgM и IgD. Мантия большинства вторичных фолликулов со стороны капсулы лимфатического узла имеет утолщение (корону). Поскольку фолликулярные дендритные клетки обладают способностью длительно удерживать на своей поверхности антиген, считают, что это способствует пролиферации расположенных рядом В-лимфоцитов, формированию центров размножения В-лимфоцитов, в центрах размножения появляются условия для образования клеток памяти. Клетки памяти остаются в зоне мантии, другие дифференцирующиеся В-лимфоциты или пополняют рециркулирующий клеточный пул или мигрируют в медуллярные тяжи. Таким образом, как отмечалось выше, формирование антителообразующих клеток пространственно отделено от участков антигенной стимуляции клеток. Эта закономерность характерна не только для лимфатических узлов, но и для селезенки.
Локализация T- и В-лимфоцитов в лимфатическом узле
Как отмечалось выше, наружные слои коры лимфатических узлов (кортикальная зона) с первичными фолликулами являются зоной локализации В-лимфоцитов. В пространствах паренхимы лимфатического узла, окружающих фолликулы, содержатся не только B-, но и Т-лимфоциты. Смесь В- и Т-лимфоцитов содержится также в мозговом слое узла. Паренхима этой области характеризуется образованием мякотных тяжей, идущих от внутренних областей кортикальной зоны к воротам узла и включающих ретикулиновые волокна, T- и В-лимфоциты с преобладанием В-клеток над Т-клетками, макрофаги и плазматические клетки, количество которых существенно возрастает в период формирования иммунного ответа.
Наибольшее количество Т-клеток регистрируется в паракортикальной зоне. Эта область содержит исключительно Т-лимфоциты, отличается наличием посткапиллярных венул с высоком эндотелием, способствующих диапедезу Т-лимфоцитов через стенку капилляров и их эмиграции в кровоток. В процессе развития клеточного иммунитета эта зона существенно увеличивается, под влиянием тимэктомии она атрофируется. В связи с этим эта зона именуется Т-зависимой областью. Помимо Т-лимфоцитов Т-зависимая область содержит большое количество интердигитальных дендритных клеток с длинными пальцевидными отростками, впервые описанных D. Veldman и отличающихся по происхождению и функциям от фолликулярных дендритных клеток. Макрофаги локализуются в различных областях узла, особенно в местах расположения T- и В-лимфоцитов. Большое количество макрофагов располагается в стенке краевого синуса и в лимфатических протоках мозгового слоя.
По строению, но не по функциям, лимфатические узлы могут различаться у разных видов животных. Так, у домашних свиней, лесных африканских кабанов, бегемотов, бородавочников, носорогов, дельфинов лимфатический узел состоит из нескольких узелков. Корковая область каждого из узелков ориентирована к центру, окружена паратрабекулярным лимфатическим синусом, вокруг которого располагается паракортикальная зона. Медуллярная (мозговая) область узла является общей для всех узелков и изолирует паракортикальную зону каждого из них. Каждый узелок омывается лимфой соответствующего ему афферентного лимфатического сосуда. Лимфа проникает в глубь узла через паратрабекулярные синусы и вытекает из него через единый эфферентный лимфатический сосуд. Мозговая область узла содержит крайне малое количество синусов, плотно упакована ретикулярными клетками, относительно недоступна для циркуляции лимфоцитов через лимфу. Вследствие этого через мозговой слой узла в лимфу эмигрирует очень небольшое количество лимфоцитов. По структуре корковая и паракортикальная области узла не отличаются от таковых других млекопитающих.
В отличие от селезенки или пейеровых бляшек, где преобладают В-лимфоциты, в лимфатическом узле преобладают Т-лимфоциты. В среднем их больше, по сравнению с В-лимфоцитами, на 20%. Под влиянием антигенного стимула в лимфатическом узле развивается как гуморальный, так и клеточный иммунный ответы. Тем не менее, преобладающей формой ответа узла на антиген являются реакции клеточного типа. Это находит объяснение в условиях органа, способствующих дифференцировке Т-лимфоцитов фенотипа CD4 в направлении Т-хелперов типа Th1, обеспечивающих формирование клеточного иммунитета.
- Сумка фабрициуса птиц
- Тимус (вилочковая железа)
- Костный мозг
- Органо-циркуляторная организация системы иммунитета
- Периферические органы системы иммунитета
- Центральные органы системы иммунитета
- Открытия в области иммунологии, удостоенные Нобелевской премии
- Развитие иммунология в России
- Аллергология
- Теории иммунитета
Изучение лимфатической системы и топографии лимфатических узлов у различных видов сельскохозяйственных животных по атласам и методическим рекомендациям.
Место проведения занятия – лаборатория ветсанэкспертизы, самостоятельная работа по атласам, учебникам и методическим рекомендациям.
Задания и вопросы для проведения текущей аттестации по первому разделу (этапу) практики.
Задание №1: изучить топографию лимфатических узлов головы жвачных животных и свиней.
Задание №2: изучить топографию лимфоузлов внутренних органов жвачных животных и свиней.
Задание №3: изучить топографию лимфатических узлов туши жвачных животных и свиней.
Задание №4: изученные лимфатические узлы крупного рогатого скота, свиней, записывают в отчете по практике в следующую таблицу.
Вопросы:
1. Значение исследования лимфатической системы для ветеринарно-санитарной экспертизы.
2. Схема лимфообращения, строение и топография лимфатических узлов и их особенности у различных видов животных.
3. Анатомо-топографические особенности лимфоузлов кролика, нутрии, домашней птицы.
5. Изменения лимфатических узлов.
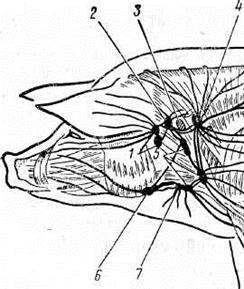 Лимфатические узлы головы свиньи.
Лимфатические узлы головы свиньи.
1 — околоушный узел.
2 — заглоточный латеральный у.,
3 — заглоточный медиальный у.,
4 — шейный поверхностный дорсальный у.,
5 — шейный поверхностный вентральный у.,
6 — подчелюстной у.,
7 — подчелюстной добавочный у.,
Рис. 1
1 — Околоушные лимфатические узлы- ln. Parotideus. Расположены под передним краем околоушной слюнной железы ниже челюстного сустава и сзади ветви нижней челюсти. Лимфа поступает из кожи головы, костей и мышц головы, жевательных мышц, ушной раковины, передней части носовой полости, околоушной и подчелюстной слюнных желез. Оттекает лимфа в заглоточные латеральные лимфатические узлы.
2 — Заглоточный латеральный лимфатический узел- ln. retropharingeus lateralis. Находится позади околоушных, прикрыты верхним концом околоушной слюнной железы. Лимфа поступает из кожи задней половины головы, ушной раковины, околоушной слюнной железы, глотки, гортани, неба, миндалин, костей мозгового черепа, околоушных лимфатических узлов. Оттекает лимфа в поверхностные шейные дорсальные и вентральные лимфатические узлы, иногда в заглоточный медиальный узел.
3 — Заглоточный медиальный лимфатический узел- ln. Retropharingeus medialis. Лежит на дорсолатеральной стенке глотки, за подъязычной костью. Собирает лимфу из жевательных мышц, сгибателей головы, костей основания черепа, задней части носовой полости, неба, глотки, миндалин; отдает лимфу в трахеальный ствол.
Поверхностные шейные лимфатические узлы делятся на дорсальные и вентральные.
4 — Шейный поверхностный дорсальный лимфатический узел- ln. cervicales superficiales dorsales. Дорсальные поверхностные шейные узлы длиной 4,5 см, всего 1—3, располагаются впереди плечевого сустава под плечеатлантной и трапециевидной мышцами. Лимфа поступает из кожи; заглоточных латеральных и вентральных поверхностных шейных узлов; дорсальной половины шеи и грудной клетки; грудной конечности. Оттекает лимфа в трахеальный проток или непосредственно в яремную вену.
5 — Шейный поверхностный вентральный лимфатический узел- ln. cervicales superficiales ventrales. Вентральные поверхностные шейные лимфатические узлы 3 — 8, лежат в яремном желобе от плечевого сустава до околоушной слюнной железы. 1 — 3 узла находятся на лестничной мышце. Собирают лимфу из нижней половины шеи и грудных стенок; части брюшных стенок; части грудной конечности; подчелюстных и околоушных лимфатических узлов. Отток лимфы происходит в дорсальные поверхностные шейные, частично в задние шейные глубокие узлы или в трахеальный ствол.
6 — Подчелюстные лимфатические узлы- ln. Mandibularis.Лежат между челюстями впереди подчелюстной слюнной железы, прикрыты передним углом околоушной слюнной железы. Собирают лимфу из передней половины головы, жевательных мышц, языка, миндалин, слюнных желез, гортани. Отток лимфы идет в поверхностные шейные вентральные, в добавочные подчелюстные и заглоточные латеральные лимфатические узлы.
7 — Добавочные подчелюстные лимфатические узлы (у свиньи) — ln. mandibularis accessorii — в количестве 2-3, величиной 0,3-1 см, лежат медиально от каудо-вентрального угла околоушной слюнной железы, на каудальном конце подчелюстной слюнной железы, у места деления яремной вены (у взрослых свиней имеются не всегда). Корни: околоушная область и передняя половина шеи. Отток лимфы идет в поверхностные шейные и заглоточный латеральный лимфоузлы.
Лимфатические узлы головы крупного рогатого скота.
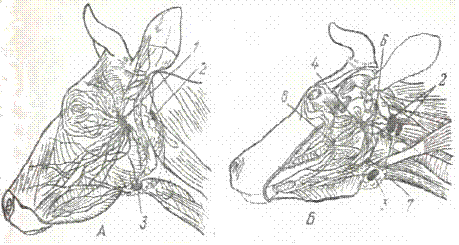
Рис. 2. Лимфатические узлы головы:
А и Б — коровы (поверхностные и глубокие); 1 — околоушной; 2 — заглоточный латеральный; 3 — подчелюстной; 4 — заглоточный медиальный; 5 — краниальные шейные; 6 — аборальный и 7 — оральный подъязычные; 8 — крыловой;
Околоушный лимфатический узел — lymphonodus parotidicus парный, длиной 6 — 9 см, вытянутой формы, лежит вентрально от челюстного сустава, задняя половина прикрыта околоушной слюнной железой. Собирает лимфу с кожи головы, с большей части мышц и костей головы, с передней половины носовой полости, губ, десен, наружного уха. Отток лимфы происходит в заглоточный латеральный лимфатический узел.
Заглоточный латеральный лимфатический узел — lymphonodus retropharyngicus lateralis парный, длиной 4 — 5 см лежит спереди и сбоку крыла атланта под задним краем околоушной слюнной железы. Собирает лимфу с кожи задней части головы и начала шеи, с губ и щек, с жевательных мускулов, с нижней челюсти, наружного уха и ушных мускулов, со многих мускулов шеи; воспринимает лимфу из всех лимфатических узлов головы. Лимфа оттекает в трахеальные протоки соответствующей стороны шеи.
Боковые заглоточные лимфатические узлы являются центральным местом сбора всех лимфатических сосудов головы и отчасти передней половины шеи. К сожалению, нередко эти узлы при отделении головы отходят к туше или разрушаются. Для их сохранения необходимо следить за тем, чтобы голова отделялась от туши на линии, проходящей между третьим и четвертым кольцом трахеи.
Подчелюстной лимфатический узел -lymphonodus mandibularis парный, длиной
2,0 — 4,5 см, овальной формы, расположен позади сосудистой вырезки нижней челюсти в межчелюстном пространстве, латеральнее подчелюстной слюнной железы. Собирает лимфу с кожи головы, с большей части мышц и костей головы, с губ и щек, передней половины носовой полости, с десен и твердого неба, с передней части языка. Лимфа оттекает в заглоточный латеральный лимфатический узел.
Заглоточный медиальный лимфатический узел (рис.2). — lymphonodus retropharyngicus medialis парный, продолговатой формы, длиной от 3 — 6 см лежит на мускулатуре глотки медиально от большой ветви подъязичной кости или между глоткой и сгибателями головы. Правый и левый узлы находятся рядом. Собирает лимфу с кожи задней части головы и начала шеи, со всего языка, с твердого и мягкого неба, миндалин, задней половины носовой полости и придаточных пазух, с глотки и гортани. Отток лимфы идет в заглоточный латеральный узел.
Ряд лимфатических узлов встречается непостоянно; к ним относятся крыловой, оральный подъязычный и аборальный подъязычный узлы.
Крыловой лимфатический узел — lymphonodus pterygoideus (рис. 2), длиной 0,75 — 1,0 см лежит позади бугра верхней челюсти, сбоку крылового мускула. Собирает лимфу из твердого и мягкого неба, десен. Отток лимфы идет в подчелюстной лимфатический узел.
Оральный подъязычный лимфатический узел- lymphonodus hvoideus oralis, расположен латерально на больших рогах подъязычной кости. Лимфа поступает из языка, а отток происходит в заглоточные узлы.
Аборальный подъязычный лимфатический узел — lymphonodus hvoideus aboralis, находится сбоку от верхнего конца среднего членика подъязычной кости. Собирает лимфу из нижней челюсти, а отток происходит в заглоточный латеральный узел.
>Энциклопедия владельца птицы
Сердечно-сосудистая и лимфатическая системы птиц
К органам крово-лимфообращения относятся сердце, кровеносные и лимфатические сосуды, кровь и лимфа. В кровеносной системе протекает кровь, приводимая в движение сердцем и стенками кровеносных сосудов, в лимфатической — лимфа, протекающая по лимфатическим протокам, лимфоузлам и вливающаяся в общий кровоток. Функции этих систем взаимосвязаны.
Кровообращение у птиц разделено на два потока артериальной и венозной крови, что обусловлено строением сердца и артериальных дуг.
Сердце птиц
Сердце является центральным органом кровообращения. Оно осуществляет циркуляцию крови по замкнутой системе кровеносных сосудов.
Сердце — полостной мышечный орган конусовидной формы, который заключен в околосердечную сумку (перикард) и вместе с ней свободно размещается в грудной полости. У птиц сердце расположено несколько правее, чем у большинства млекопитающих, и частично прикрыто воздухоносными мешками. Верхушка сердца расположена между долями печени и мышечным отделом желудка. Форма сердца чаще коническая, а у некоторых птиц — сильно удлиненная. У разных видов птиц форма сердца может значительно отличаться. Оба желудочка внешне мало разграничены, хотя и имеются слабо выраженные вентральная (со стороны живота) и дорсальная (со стороны спины) межжелудочковые борозды.
Стенка сердца состоит из мышечного слоя — миокарда. Изнутри полость сердца выстлана тонкой оболочкой — эндокардом, а снаружи покрыта серозной оболочкой — эпикардом.
Внутренняя полость сердца разделена продольной перегородкой на правую и левую половины. Каждая из них, в свою очередь, разделена на две камеры. Всего в сердце имеется четыре камеры: две верхние — левое и правое предсердия, две нижние — левый и правый желудочки.
Левое предсердие принимает, а левый желудочек выталкивает артериальную кровь, насыщенную кислородом. Правое предсердие принимает, а правый желудочек выталкивает венозную кровь, насыщенную углекислотой. На поверхности сердца имеются две бороздки: одна из них поперечная, на границе между предсердиями и желудочками, а другая — продольная, на границе между желудочками.
В левое предсердие впадают правая и левая легочные вены, которые прямо перед сердцем сливаются в один ствол, переходящий в предсердие.
В правое предсердие впадают две передние и одна задняя полые вены. Они снабжены перепончатыми клапанами, которые препятствуют обратному поступлению крови из предсердия в вены. В перегородке предсердий имеется овальная впадина — остаток овального отверстия, имеющегося в сердце эмбриона.
Снабжение кровью самого сердца осуществляется по двум коронарным артериям, отток венозной крови — по трем венам. Степень капилляризации миокарда тесно связана с величиной сердца. Сердце у птиц относительно велико и по отношению к массе тела значительно превосходит сердце у млекопитающих. Например, у колибри его масса составляет до 2,75% массы всего организма. Масса сердца у мелких птиц относительно большая, чем у крупных, что связано с более интенсивным обменом веществ (в свою очередь это обусловлено тем, что у мелких животных менее выгодное соотношение между объемом тела и поверхностью, отдающей тепло). Например, у снегиря при массе тела в 23 г относительная масса сердца 1,3%, у чечетки массой 13 г – 1,6%, а у синицы московки массой 8 г – 1,8%.
Существует также зависимость между относительной массой сердца и энергичностью движений. У хорошего летуна сокола чеглока масса сердца составляет в среднем 1,7% от массы тела, у менее хорошего летуна пустельги – 1,2%, а у плохого летуна сороки – всего около 0,9%. У всех птиц, которые часто летают, сердце должно быть крупным, чтобы обеспечивать быструю циркуляцию крови. То же самое можно сказать о видах, обитающих в холодных областях или на больших высотах.
У теплокровных сердечный индекс (уровень кровоснабжения самого сердца кровью) в несколько раз больше, чем у холоднокровных. Сердечный индекс у птиц около или выше 10% (сокол — 15,57%, воробей — 13,53%, дикая утка — 9,85%). Большая вариабельность наблюдается у млекопитающих, но в общем они имеют высокий сердечный индекс, хотя и меньший, чем птицы. Соответственно, наибольшее количество капилляров в 1 мм² миокарда отмечается у птиц, обладающих высоким сердечным индексом: у орла, голубя, чайки, вороны. У домашних нелетающих птиц этот показатель несколько ниже. Большая ширина капилляров миокарда птиц по сравнению с млекопитающими связана с более крупными эритроцитами. По интенсивности кровоснабжения сердце птиц превосходит сердце млекопитающих животных, и его кровоснабжение соответствует высокоорганизованному сердцу (см. таблицу 1). Это обусловлено большой функциональной нагрузкой, которую выполняет сердце птиц во время полета.
Таблица 1. Степень капилляризации миокарда и диаметр капилляров у разных классов животных
| Животные | Количество капилляров в 1мм2 | Диаметр капилляров, мкм |
| Рептилии | 1276 | 15 |
| Птицы | 4047 | 8,5 |
| Млекопитающие | 3512 | 6,5 |
С эволюционной точки зрения, сердце птиц занимает одну из верхних ступеней развития, что в свою очередь имеет решающее значение для эволюции различных органов.
В течение всей жизни сердце работает ритмично и обеспечивает циркуляцию крови в кровеносной системе. Ритмичная работа сердца достигается тем, что в нем имеется особая нервно-мышечная система, проводящая возбуждение. В составе сердечной мышцы есть нервно-мышечные образования, которые отличаются от волокон типичной мускулатуры сердца; они менее исчерчены и содержат меньше белковых нитевидных структур, с помощью которых происходит мышечное сокращение. Эти специфические мышечные образования и составляют нервно-мышечную систему сердца.
Мышечные ткани сердца эмбриона птиц еще до прорастания в них нервов начинают ритмично сокращаться. Способность сердца сокращаться ритмично независимо от внешних раздражителей называется автоматизмом. Автоматизм сердца обусловлен наличием нервно-мышечной проводящей системы.
Работа сердца у птиц более энергична, чем у низкоорганизованных наземных позвоночных. У травяной лягушки число сокращений сердца в минуту 40-50, у снегиря – в среднем 730. Хорошо видна зависимость частоты сокращений сердца от размеров тела. У голубя массой 250 г число сокращений сердца в минуту в среднем 248, у зеленушки массой 22 г – 697, у щегла массой 13 г – 754, у синицы московки массой 8 г – 1037. Частота сердечных сокращений зависит и от состояния птицы, от вида и возраста, функционального состояния мышц сердца и нервных возбуждений, к ним поступающих, температуры окружающего воздуха. Например, сердце взрослых птиц может делать 200 – 300, а молодняка того же вида птиц – 400 – 500 сокращений в минуту, или у голубя в покое она равна в среднем 165 ударам в минуту, а в полете – 550. У канарейки массой 16 г, содержащейся в домашних условиях частота сердечных сокращений (ЧСС) доходит до 1000 ударов, хотя ее масса в 2 раза больше, чем у синицы московки, живущей в естественных условиях (ЧСС – 1037). У водоплавающих и ныряющих птиц при нырянии возникает явление брадикардии (временного замедления ритма). Так, у оляпки (ныряющей птицы) через 5 с после погружения в воду сердечный ритм уменьшается от первоначального до 73%, через 10 с – до 48% и через 15 с – до 42%.
Деятельность сердца состоит из трех фаз. В первую фазу сокращаются левое и правое предсердия, желудочки расслабляются (отдыхают). Во вторую фазу сокращаются левый и правый желудочки, а предсердия расслабляются (отдыхают). Третья фаза (общая диастола) – одновременно расслабляются предсердия и желудочки (фаза отдыха). Последовательное сокращение предсердий и желудочков называется систолой, расслабление мышц сердца – диастолой. Три фазы деятельности составляют сердечный цикл. Необходимо отметить, что суммарная фаза отдыха, как предсердий, так и желудочков сердца больше фазы работы, поэтому сердце никогда не устает. Такой сердечный цикл свойственен не только птицам, но и всем млекопитающим. При крайнем нервном возбуждении или сильном заболевании возможна значительная тахикардия (резкое и длительное увеличение ЧСС). В таких случаях появляется риск износа мышечной ткани сердца за счет недостаточного кровоснабжения самой сердечной мышцы. Это состояние может привести к инфаркту миокарда.
Большой и малый круги кровообращения. Строение сосудов
Циркуляция крови в организме происходит по замкнутой системе кровеносных сосудов. Артерии, вены, капилляры, сердце, лимфатические протоки и лимфоузлы составляют единую замкнутую сердечно-сосудистую систему. Кровеносную систему делят на два круга кровообращения — большой и малый.
Артериальная кровь из левого желудочка попадает в аорту, которая затем делится на ряд артерий. Таким образом, артериальной кровью снабжаются голова, пояс передних и задних конечностей, сами конечности, мышцы брюшной стенки, органы таза, желудок и кишечник. После снабжения кислородом всех органов и тканей, уже венозная кровь поступает в вены, которые собираются в две большие полые вены, и впадает в правое предсердие, а затем в правый желудочек. Это составляет большой круг кровообращения.
Малый круг кровообращения начинается движением крови из правого желудочка, далее по общей легочной артерии, несущей к легким венозную кровь. После прохождения по системе легочных капилляров и уже насыщенная кислородом кровь собирается в вену и поступает в левое предсердие, а далее в левый желудочек, где заканчивается малый круг кровообращения.
Кровеносные сосуды проводят кровь и способствуют ее продвижению от сердца к тканям и обратно, регулируют кровенаполнение органов тела в зависимости от функции последних, изолируют кровь от непосредственного соприкосновения с тканями.
По строению и выполняемой функции кровеносные сосуды разделяются на проводящие (артерии, вены, артериовенозные анастомозы) и питающие (капилляры). Строение кровеносных сосудов соответствует выполняемой функции. Наибольший диаметр (0.5 — 1см) имеют крупные артерии, отходящие от сердца. По мере удаления от сердца диаметр их уменьшается, и они постепенно переходят в артериолы, а последние — в прекапилляры (диаметр 15-20 мкм) и затем в капилляры (диаметр просвета капилляра соответствует толщине одного эритроцита); далее капилляры артерий переходят в посткапилляры, а последние — в венулы (диаметр 30-40 мкм). Сливаясь и укрупняясь, венулы образуют вены. Комплекс этих сосудов называется микроциркуляторным руслом, а процесс движения крови по ним – микроциркуляцией. От нее в значительной степени зависят адаптация организма к условиям среды и уровень обменных процессов, жизнь клеток и всего организма. Так, в прекапиллярах имеются сфинктеры; расслабляясь, они пропускают кровь в капилляры, а сокращаясь, препятствуют ее движению. Это необходимо для регулирования кровотока при переходе из состояния покоя в состояние физической активности. В состоянии покоя довольно большая часть прекапилляров закрыта, а в состоянии физической нагрузки практически все прекапилляры открыты. Также это имеет значение при перераспределении кровотока между органами и тканями. В зависимости от физической нагрузки и уровня обмена веществ в организме количество капилляров, участвующих в кровоснабжении, может изменяться в пределах 10-90%.
Артерии несут кровь в направлении от сердца, вены – к сердцу.
Стенка вен и артерий состоит из трех слоев: наружного соединительно-тканного, среднего гладкомышечного и внутреннего, образованного одним слоем плоских клеток. Стенки артерий имеют наибольшую толщину гладкомышечного слоя, стенки вен имеют гораздо менее выраженный слой из гладких мышц. Наибольшую толщину гладкомышечного слоя имеют аорта и легочная артерия. Это обусловлено тем, что стенки этих артерий постоянно испытывают давление ударной волны крови, поступающей из сердца при его сокращении. Внутренняя стенка вен образует карманоподобные клапаны, расположенные по направлению тока крови и способствующие ее продвижению к сердцу.
Капилляры — очень тонкие кровеносные сосуды, диаметром 5-7 мкм. Они имеются в огромных количествах во всех органах и образуют густую сеть. Стенки большинства капилляров состоят только из эндотелия (одного слоя клеток), через который осуществляется тканевой и межуточный обмен веществ. Через эндотелий капилляров в межклеточные пространства тканей проникают плазма крови и содержащиеся в ней молекулы белков, жиров, углеводов. Также питательные вещества и жидкость проникают в клетки через поры клеточных мембран. Через капилляры происходит диффузия кислорода, углекислого газа и других веществ.
Физиология кровеносных сосудов и движение крови по ним
Непрерывное движение крови в сосудах обусловлено деятельностью сердца и разностью давления на артериальных и венозных концах сосудистой системы. В передвижении крови по организму активное участие принимают и сами кровеносные сосуды, а продвижению крови по венам помогают скелетные мышцы. При сокращении мышц кровь в венах продвигается только к сердцу, так как клапаны в венах открываются лишь в сторону сердца.
В момент сокращения сердечных мышц полости сердца сужаются, и давление в них повышается, кровь под давлением выбрасывается в аорту. В момент расслабления создается разреженное пространство в полостях сердца, кровь притекает по венам и заполняет полости.
Скорость течения крови зависит от суммарной емкости кровеносных сосудов. Там, где суммарная емкость сосудов маленькая, скорость течения крови большая. Например, в аорте скорость течения в среднем составляет 0.6 м/с, а в момент систолы до 1 м/с; в капиллярах суммарная емкость самая большая, поэтому скорость течения крови в них самая меньшая (от 0.3 до 0.16 мкм/с). Замедленное течение крови в капиллярах способствует лучшему обмену веществ между кровью и тканевой жидкостью.
Стенки сосудов эластичны и под давлением крови, поступающей в результате сокращения сердца, растягиваются в период систолы и спадаются в период диастолы. При этом они давят на кровь и проталкивают ее вперед, создавая кровяное давление. В зависимости от эластичности сосудов и величины их просвета кровяное давление может быть различным. Например, у петуха в сонной артерии оно равно 200 мм ртутного столба, в бедренной – 140 мм ртутного столба; у индейки в бедренной артерии кровяное давление колеблется между 78 – 122, у индюка – 160 – 170 мм ртутного столба, у голубя оно равно – 135/105. Кровяное давление в норме поддерживается на постоянном уровне. Крайний испуг может настолько повысить у птиц кровяное давление, что крупные артерии лопаются и особь умирает.
Уровень артериального давления птиц также является подтверждением высокого уровня метаболизма. Разницу в уровне артериального давления у разных классов животных можно увидеть в таблице 3.
Таблица 2. Показатели артериального давления у различных классов животных
| Класс животных | Артериальное давление мм рт. ст. | |
| Среднее | Высшее | |
| Рептилии | 25 | 45 |
| Птицы | 120 | 230 |
| Млекопитающие | 100 | 150 |
Примечание: 1 мм рт. ст. равняется 0,1333 кПа.
Дополнительно у птиц существует система регулирования ширины просвета сосудов. Регуляция просвета сосудов осуществляется при участии нервов. В продолговатом мозге имеется сосудодвигательный центр – скопление двух групп клеток; одна из них регулирует расширение, другая – сужение сосудов. В спинном мозге есть также сосудодвигательные центры, но они проявляют самостоятельное влияние только в том случае, когда нарушается регулирующее действие со стороны вышележащих отделов центральной нервной системы. Нервы, изменяющие просвет сосудов, называются сосудодвигательными. Они могут быть сосудосуживающими и сосудорасширяющими. Сосудосуживающие (вазоконстрикторы) – это нервы симпатического отдела нервной системы, тела этих нервных клеток заложены в спинном мозге. Центры сосудорасширяющих нервов (вазодиляторы) расположены в межпозвоночном узле. Сосудорасширяющими нервами в основном являются афферентные (чувствительные) волокна, которые выполняют двойную функцию: проводят возбуждение к спинному мозгу (это их прямое назначение) и приносят импульсы в обратном направлении, от центра к мышцам стенок сосудов. На просвет кровеносных сосудов могут оказывать влияние и гуморальные факторы – гормоны и некоторые продукты обмена веществ. Обычно в организме стенки артерий несколько напряжены и просвет их сужен. Это состояние постоянного напряжения называется сосудистым тонусом.
Терморегуляция у птиц
В связи с разделением большого и малого кругов кровообращения все органы омываются чистой артериальной кровью. Это обстоятельство, а также быстрая циркуляция крови и энергично идущий газообмен обусловливают высокую температуру тела, в среднем равную + 42° С. У крупных птиц она обычно +38°…+45° С.
У птиц существует как химическая, так и физическая терморегуляция. Первая состоит в изменении интенсивности обмена, т.е. величины теплопродукции в зависимости от температуры внешней среды, количества и качества потребляемой пищи. Так, падение внешней температуры с +33° до +10°С вызывает у воробья повышение потребления кислорода в три раза. Физическая терморегуляция заключается в изменении величины теплоотдачи. У птиц существенное значение имеет так называемая тепловая одышка, заключающаяся в учащении дыхания, что приводит к увеличению отдачи тепла с выдыхаемым воздухом и испарению влаги из органов дыхания и дыхательных путей. Этим способом, например, мелкие птицы могут рассеять около половины тепла, выделяемого организмом. У крупных птиц рассеивание тепла таким образом может даже превосходить теплопродукцию. Благодаря этому, например, страусы и даже голуби могут выдерживать почти без перегревания температуру среды в +50°С.
К системе органов лимфообращения относятся лимфатические пространства, капилляры, лимфатические сосуды, лимфатические узлы и лимфа. Лимфа движется по капиллярам и лимфатическим сосудам. Капилляры лимфатической системы — это мельчайшие просветы между клетками, они выстланы эндотелием. Капилляры впадают в лимфатические сосуды среднего и большого диаметра, которые имеют карманообразные клапаны, открывающиеся только по течению лимфы. Лимфатические сосуды всего тела соединяются в два главных ствола — левый и правый млечные, так называемые грудные протоки. Они идут краниально (по направлению к голове) по обеим сторонам позвоночного столба. Оба ствола соединяются между собой поперечными протоками и впадают в конечные отделы левой и правой яремных вен (по яремным венам отводится кровь от головы, шеи, пищевода и трахеи). Лимфатические сосуды, подходя к лимфоузлам, распадаются на сеть мельчайших сосудов, которые потом снова соединяются в крупный сосуд.
Очень важной является лимфатическая микроциркуляция, т.е. движение лимфы по лимфатическим прекапиллярам, капиллярам и посткапиллярам. Благодаря лимфатической микроциркуляции из тканей удаляются отработанные вещества, неиспользованные крупномолекулярные белки, липиды. Микроциркуляция крови и лимфы едина.
Лимфоузлы у птиц (их два) не имеют четкого разделения на мозговой и корковый слои. Узлы хорошо выражены у гусей и уток в нижней части шеи (около яремной вены) и в области поясницы (на уровне половых желез, между аортой и внутренним краем почек). Лимфоузлы имеют серовато-белый или темно-серый цвет. У кур нет типичных лимфоузлов. У них по всему телу разбросана лимфоидная ткань в виде одиночных лимфоидных скоплений (узелков без капсулы). Наиболее выраженные места лимфоидных скоплений находятся в печени, коже, стенках кишечника, легких, глотке, небе. В них, так же как и в лимфоузлах уток и гусей, есть реактивные центры.
Кровь и лимфа
Кровь — ткань внутренней среды защитно-трофической функции, состоящая из жидкого межклеточного вещества (плазмы), эритроцитов, тромбоцитов и клеток на всех стадиях своего развития в кроветворных органах. Клеточные структуры периферической крови называются форменными элементами. Эритроциты птиц, в отличие от человека, имеют ядра.
Кровь выполняет следующие функции: 1) трофическую — перенос питательных веществ ко всем клеткам и тканям; 2) дыхательную — газообменную, или транспорт кислорода к тканям и удаление из организма углекислоты; 3) защитную (фагоцитоз, выработка антител); 4) регуляторную — транспорт гормонов и других гуморальных факторов регуляции; 5) гомеостатическую — поддержание физико-химического постоянства состава внутренней среды организма. Плазма крови — это жидкое межклеточное вещество, в котором во взвешенном состоянии находятся форменные элементы крови. Основу плазмы составляет вода, остальное — белки (альбумины, глобулины, фибриноген и десятки других), липиды, углеводы, минеральные вещества.
Прогрессивной чертой птиц является и общее увеличение объема крови. У костистых рыб масса крови составляет примерно 3% от массы тела, у бесхвостых амфибий – 6%, у птиц – 9%. Кислородная емкость крови у птиц примерно в 2 раза большая, чем у рептилий. Все эти черты организации являются важной предпосылкой общего повышения жизнедеятельности птиц. Также кровь птиц обычно содержит больше эритроцитов, чем у большинства млекопитающих, и в результате в единицу времени может переносить больше кислорода, что необходимо для полета.
Лимфа — бесцветная мутноватая жидкость, по своему химическому составу и осмотическому давлению напоминает плазму крови. Она свертывается, в ее составе есть белок фибриноген. Воды в лимфе содержится 94-95%, сухого остатка — 5-6%. Лимфа заполняет лимфатические протоки, а также содержится (в виде серозной жидкости) в различных полостях тела (желудочек мозга, полости суставов, сердечной сумки, плевры, брюшины).
Некоторые особенности биохимии крови попугаев
Систематические исследования биохимических показателей крови здоровых попугаев появились сравнительно недавно. Имеются значительные отличия в биохимических показателях плазмы между родами отряда Попугаи и поэтому нельзя дать усредненные значения для отряда в целом. Интерпретация результатов биохимии крови определенного вида попугаев может проводиться, если нормальные значения для данного вида установлены теми же самыми методами.
Содержание общего белка в крови попугаев возможно не столь показательно, как соотношение отдельных электрофоретических фракций, однако уровень общего белка в целом неспецифически характеризует состояние птицы и резкие отклонения от нормы отражают грубые обменные патологии. Было определено, что концентрация общего белка в норме незначительно варьирует у представителей разных родов.
По результатам исследований выявлено, что у попугаев всех исследованных таксономических групп с возрастом снижается уровень мочевины и повышается уровень мочевой кислоты в крови. Оба эти параметра отражают ход естественной биодеградации азотсодержащих молекул (нуклеиновые кислоты, белки) и их следует рассматривать в комплексе. Крупные попугаи (ара, какаду) имеют более низкие уровни и мочевины и мочевой кислоты, чем мелкие виды (карелла) что, возможно, связано с большей интенсивностью у последних обменных процессов.
В норме концентрация глюкозы в крови отличается у представителей разных родов. Так например, у жако она составляет около 12 ммоль/л, а у ары — около 7 ммоль/л. Это необходимо учитывать. У молодых попугаев уровни глюкозы выше. Увеличение содержания глюкозы происходит при сахарном диабете, который описан у карелл, амазонов, какаду и ары. Уменьшение — при дистрофиях, голодании, дисфункциях печени, сепсисе и новообразованиях.
Нормальные значения холестерина варьируют между родами, хотя рацион у попугаев примерно одинаков. Так, у какаду холестерина на 50% больше, чем у ары.
Вариации концентрации кальция в крови попугаев незначительны, т.к. кальций одинаково важен для представителей всех родов попугаев. Ионы кальция участвуют в акте мышечного сокращения, проведении нервного импульса, процессах свертывания крови и входят в состав костной ткани. Не удивительно, что у молодых птиц уровень кальция в крови ниже, чем у взрослых т.к. в молодом организме процессы перераспределения кальция более интенсивны. Например, у карелл он составляет 2,4 и 3,0 ммоль/л. Многие авторы описывают гипокальциемию у жако. Для этого рода попугаев особенно важно знать нормальный уровень кальция в крови, т.к. у других попугаев гипокальциемия не встречается.
Список литературы и интернет источников
1. Кульчицкий К.И., Роменский О.Ю. «Сравнительная анатомия и эволюция кровеносных сосудов сердца» — К.: «Здоровья», 1985. — 176с.
2. «Зоология позвоночных: учебник для студентов высших учебных заведений/ Константинов В.М., Шаталова С.П. – М.: Гуманитар. изд. центр ВЛАДОС, 2004. – 527 с.: ил.»
3. Константинов В.М., Шаталова С.П. Сравнительная анатомия позвоночных животных. М., 2005.
4. Наумов Н.П., Карташов Н.Н. Зоология позвоночных. Ч.2: Пресмыкающиеся, Птицы, Млекопитающие. М., 1979.
5. Мамонтов С.Г. Биология. Для школьников старших классов и поступающих в вузы: Учеб. пособие. – 2-е изд. – М.: Дрофа, 1999. – 480 с.: ил.
6.
7.
8.
9.
Ссылки на рисунки:
Лимфатическая система
К системе органов лимфообращения относятся лимфатические пространства, капилляры, лимфатические сосуды, лимфатические узлы и лимфа.
Лимфа – бесцветная мутноватая жидкость, по своему химическому составу и осмотическому давлению напоминает плазму крови. Она свертывается, в ее составе есть белок фибриноген. Воды в лимфе содержится 94-95%, сухого остатка – 5-6%. Лимфа заполняет лимфатические протоки, а также содержится (в виде серозной жидкости) в различных полостях тела (желудочек мозга, полости суставов, сердечной сумки, плевры, брюшины).
Образование лимфы. Есть две теории образования лимфы. Согласно физико-химической, или фильтрационной, теории, через стенки капилляров в межклеточные пространства просачивается жидкая часть крови, а эритроциты и лейкоциты задерживаются в кровеносных сосудах. Таким путем образуется межклеточная, или тканевая, жидкость. Она омывает все клетки. Из крови в нее поступают питательные вещества и кислород для клеток , а из нее в кровь – продукты обмена веществ. Тканевая жидкость, поступая в лимфатические протоки, дополняется лимфоцитами и становится лимфой. Секреторная теория объясняет образование лимфы тем, что ее секретируют клетки, составляющие стенки капилляров.
Передвижение лимфы. Лимфа движется по капиллярам и лимфатическим сосудам. Капилляры лимфатической системы – это мельчайшие просветы между клетками, они выстланы эндотелием. Капилляры впадают в лимфатические сосуды среднего и большого диаметра, которые имеют карманообразные клапаны, открывающиеся только по течению лимфы.
Лимфатические сосуды всего тела соединяются в два главных ствола – левый и правый млечные, так называемые грудные протоки. Они идут краниально по обеим сторонам позвоночного столба по направлению к шее. Оба ствола соединяются между собой поперечными протоками и впадают в конечные отделы левой и правой яремных вен.
Стенки лимфатических сосудов очень тонкие и легко спадаются под давлением. Но в каудальной части тела имеются расширения лимфатических сосудов со значительным мышечным слоем. Они обладают способностью сокращаться и продвигать лимфу. Эти участки получили название «лимфатических сердца». Движению лимфы способствуют сокращение мышц тела и давление наполненных воздухоносных мешков.
Лимфатические сосуды, подходя к лимфоузлам, распадаются на сеть мельчайших сосудов, которые потом снова соединяются в крупный сосуд.
Лимфоузлы у птиц (их два) не имеют четкого разделения на мозговой и корковый слои. Узлы хорошо выражены у гусей и уток в нижней части шеи (около яремной вены) и в области поясницы (на уровне половых желез, между аортой и внутренним краем почек). Лимфоузлы имеют серовато-белый или темно-серый цвет. Они состоят из соединительнотканной капсулы, вторичных лимфоузлов округлой, и многоугольной формы, расположенных по всему лимфоузлу, и небольшого числа кровеносных сосудов. Во вторичных узелках имеются так называемые реактивные центры, где размножаются лимфоциты, а по некоторым данным, и гранулоциты (Г.С.Крок). У кур нет типичных лимфоузлов. У них по всему телу разбросана лимфоидная ткань в виде одиночных лимфоидных скоплений (узелков без капсулы). Наиболее выраженные места лимфоидных скоплений находятся в печени, коже, стенках кишечника, легких, глотке, небе. В них, так же как и в лимфоузлах уток и гусей, есть реактивные центры.
Лимфатическая система птичьего эмбриона
Данных о морфологии и физиологии лимфатической системы у птичьего эмбриона очень мало.
Лимфатическая система куриного эмбриона развивается сравнительно поздно и появляется на 5—6-й дни инкубации двумя центрами: один в области таза и второй (более крупный) — у основания шеи. Они соединяются лимфатическим каналом, который образуется путем слияния в мезенхиме лимфатических полостей. Лимфатические капилляры выстланы эндотелием и заканчиваются слепо. Все лимфатические капилляры сливаются в лимфатические сосуды, которые впадают в яремный лимфатический мешок, а оттуда — в венозную систему, через грудной проток, в месте слияния подключичной и яремной вен. Лимфатические сосуды возвращают часть плазмы крови, прошедшей через стенки кровеносных сосудов и омывшей клетки тканей тела. Наиболее крупные лимфатические сосуды имеют клапаны, предотвращающие обратный ток жидкости.
Путем слияния капилляров лимфатических сплетений образуются расширения лимфатических сосудов — лимфатические мешки, которые в свою очередь превращаются в лимфатические железы. Они состоят из скоплений лимфоидной ткани, продуцирующей лимфоциты. У птиц в отличие от млекопитающих очень мало лимфатических желез. Обычно имеются две пары лимфатических желез — тазовые и яремные, но у взрослых кур последних нет.
В области пояса задних конечностей расположены два маленьких лимфатических сердца, помогающих движению лимфы по лимфатическим сосудам. В эмбриональном периоде их роль в основном заключается в дренаже лимфы из аллантоиса. У многих птиц лимфатические сердца имеются только в эмбриональном периоде. Интересно отметить, что у наиболее древних птиц — страусов — лимфатические сердца особенно хорошо развиты. У кур они функционируют только в эмбриональном и начале постэмбрионального периода. Сокращения лимфатических сердец начинаются у 6—6.5-дневных куриных эмбрионов и связаны вначале со спазматическими движениями хвоста, но у 10—13-дневных сокращения лимфатических сердец уже делаются независимыми от этих движений и происходят 4—8 раз в 1 мин.
Ток лимфы начинается с 8-го дня и независим от тока крови. Лимфа состоит из плазмы и лимфоцитов, свободно проходящих через тонкие эндотелиальные стенки лимфатических сосудов. Жидкость, проникающая в межклеточные пространства из кровеносных капилляров и клеток тканей, продукты их обмена веществ, а также питательные вещества из кишечника, поступающие через лимфатические сосуды кишечного тракта, — все это составляет плазму лимфы. Лимфа — это истинная внутренняя среда организма, так как она в отличие от крови находится не в замкнутой системе сосудов, а непосредственно соприкасается с клетками тканей.
Лимфоциты, кроме лимфатических желез, образуются также в лимфоидных органах. Одним из самых крупных лимфоидных органов является селезенка, отличающаяся от лимфатических желез тем, что она не имеет непосредственного сообщения с лимфатическими сосудами, а обильно снабжается только кровью. Селезенка возникает в виде отпочкования стенки брюшной полости. У 4-дневного эмбриона это утолщение брюшины уже заметно, а затем клетки его быстро растут, и у 6-дневного эмбриона селезенка представляет собою заметное выпячивание, находящееся над желудком. Брюшинные клетки проникают в рядом лежащую мезенхиму, к которой непосредственно примыкает вена селезенки.
Тимус, по мнению Романова, является первым органом, где начинается лейкоцитопоэз, а по количеству лейкоцитов — вторым после костного мозга. Начиная с 8-го дня инкубации в тимусе происходит лимфоцитопоэз, а также образование эозинофильных гранулоцитов и в небольшой степени — эритропоэз. По данным Аккермана и Науфа, эпителиальные клетки тимуса начинают приобретать признаки лимфоцитобластов у 7.5—8-дневных куриных эмбрионов, а на 10—11-й день инкубации лимфоцитобласты превращаются в лимфоциты. Авторы подчеркивают сходство образования лимфоцитов в тимусе с тем, как они образуются в фабрициевой сумке, что было установлено ими ранее. Фабрициева сумка, как известно, — специфический для птиц лимфоэпителиальный орган. Она возникает как слепой, отпочковавшийся вырост клоаки и состоит из медулярной части, возникающей из поверхностного эпителия, и кортикальной, происходящей из мезенхимных и эпителиальных клеток. В обеих частях фабрициевой сумки происходит лимфоцитопоэз.
Лимфоидный характер носит также и остаток желточного мешка в постэмбриональном развитии. Это слепой выступ примерно на середине тонкой кишки, и его развитие обычно обратно пропорционально развитию слепой кишки, которая также является лимфоидно-эпителиальным органом.
Если вы нашли ошибку, пожалуйста, выделите фрагмент текста и нажмите Ctrl+Enter.
Исследование поверхностных лимфатических узлов входит также в состав общего исследования. Оно имеет важное значение в диагностике некоторых заразных заболеваний, давая признаки, которые вызывают у врача подозрение относительно той или иной инфекции и побуждают его к особенно подробному исследованию в определённых направлениях. В иных случаях этого рода исследование доставляет данные, которые, как, например, гнойная инфильтрация узлов с образованием абсцесса при мыте, являются даже решающими при постановке диагноза.
Кроме того, при местных процессах, а также при локализованных инфекциях, когда микроорганизмы, оседая в лимфатических узлах соответствующих областей, тем самым обусловливают явления вторичного воспаления, эти изменения используются при диагностике основного процесса и первично поражённых органов. При небольшой затрате времени исследование лимфатических узлов оказывает иногда существенную услугу и потому не должно упускаться, несмотря на некоторые трудности.
В качестве методов исследования используют осмотр и пальпацию, лишь временами, в случае нужды, прибегая к пробному проколу и экстирпации. Небольшие размеры лимфатических узлов, их расположение в толще окутывающей их клетчатки скрывают даже поверхностные лимфатические узлы от взора наблюдателя и делают невозможным прижизненное исследование большинства из них у здоровых животных. Трудность увеличивается ещё тем, что величина лимфатических узлов у домашних животных показывает большие колебания, в зависимости от возраста и породы животного. Таким образом, этого рода исследование, как будто бы очень несложное, требует не только подготовки учащегося по топографической анатомии, но и клинических навыков, которые создаются путём опыта и продолжительного упражнения.
Из поверхностно расположенных узлов у лошадей легко прощупывают подчелюстные, сра. м.ные и железы коленной складки. В случае изменений, обусловленных некоторыми патологическими процессами, кроме того, удаётся прощупать околоушные, верхние, средние и нижние шейные, предлопа-точные, локтевые, подколенной ямки, поясничные, паховые и крестцовые узлы.
У крупного рогатого скота наиболее массивными являются подчелюстные, предлопаточные, коленной складки и выменные узлы. При туберкулёзе и некоторых других заболеваниях, вследствие инфильтрации и резкого увеличения железистой ткани, удаётся легко прощупать, кроме того, три пары шейных узлов, околоушные, ретрофарингеальные и узлы голодной ямки.
Несложная в общем методика исследования подчелюстных узлов лошади состоит в том, что, надёжно фиксировав голову животного левой рукой, положенной на спинку носа, ощупывают пальцами правой руки всё подчелюстное пространство от области гортани до подбородочного угла. Пальпацию правой железы удобнее проводить левой рукой, расположившись справа от головы животного. При пальпации левой железы изменяют положение соответствующим образом.
Особого внимания при исследовании заслуживают следующие факторы: Величина узла,, его строение, форма и свойства поверхности, консистенция, температура покрывающей его кожи, чувствительность, подвижность самого узла, и, наконец, подвижность покрывающей его кожи.
В результате исследования врач должен составить себе ясное представление о каждом из отмеченных факторов, что оказывает ему существенную услугу при оценке состояния лимфатической системы.
Из изменений необходимо отметить следующие:
Острые набухания узлов, являющиеся следствием паренхиматозных воспалений железистой ткани с серозно-клеточной их инфильтрацией (лимфадениты). При этом узлы кажутся несколько увеличенными в размерах, опухшими и болезненными при надавливании. Их дольчатое строение менее заметно, консистееция становится плотной, температура несколько повышена, подвижность ограничена, а поверхность остаётся ровной и гладкой. Набухания подчелюстных узлов у лошади наблюдаются при многих острых инфекционных заболеваниях, например, при инфлуэнце, заразном катаре верхних дыхательных путей и контагиозной плевропневмонии, инфекционной анемии, остром и подостро протекающем сапе, а также при всех острых местных процессах, связанных с истечением износа,-катарах носа, воспалениях глотки, придаточных полостей лица.
Hагноение лимфатичсских узлов с образованием абсцессов является высшей степенью острого воспаления. Обширные нагноения подчелюстных лимфатических узлов с образованием громадного количества гноя и развитием воспалительного отёка окружающей клетчатки считаются характернейшим симптомом мыта. При этим область узла резко припухает, образуя заметное выпячивание. Кожа делается напряжённой, горячей наощупь и сильно болезненной. При пальпации вначале обнаруживают плотную консистенцию, которая постепенно становится мягче и переходит в ясную флюктуацию. При этом кожа там, где давление наиболее сильно, истончается, покрывающая её шерсть выпадает, на поверхности выступают капли серозного выпота, и, наконец, абсцесс вскрывается с выделением большого количества сливкообразного гноя. Иногда переход воспалительного набухания подчелюстных узлов в нагноение наблюдают и при других заболеваниях, например, при фарингитах, тяжёлых ринитах и, редко, при сапе и туберкулёзе; однако в подобных случаях нагноения являются очень ограниченными, гноя скопляется немного, а подкожная клетчатка оказывается совершенно неизменённой.
Гиперплязия лимфатических узлов при лейкемии и псевдолейкемии крупного рогатого скота, сопровождающихся равномерным и довольно значительным увеличением объёма всех поверхностных лимфатических узлов. Подобного рода симметричные припухания иногда наблюдают у крупного рогатого скота при туберкулёзе. Однако чаще туберкулёзные поражения (лимфатических узлов) у крупного рогатого скота проявляются в форме заметных при осмотре припуханий только отдельных узлов, которые становятся плотными, бугристыми, полностью сохраняя, однако, свою подвижность. Иногда туберкулёзно изменённая железа нагнаивается с образованием густого гноя. У свиньи при туберкулёзе, проявляющемся обычно в форме поражения заглоточных, шейных или подчелюстных лимфатических узлов (скрофулёз), наблюдают резкие припухания, сначала плотные и безболезненные, которые потом, вследствие творожистого перерождения, флюктуируют и вскрываются с выделением казеозных масс. Характер изменений устанавливается прививкой ткани Железы морским свинкам, особенно чувствительным к туберкулёзной инфекции.
Хроничекая деформация лимфатически х узлов вследствие разрастания соединительной ткани железы. Хронически действующее раздражение приводит к разрастанию соединительной ткани не только в самой железе, по ипарааденонднойткаии, равно как и окружающей всю железу Подкожной клетчатке. Вследствие сморщивания соединительной ткани совершенно изменяются свойства железы; она становится плотной, бугристой, совершенно нечувствительной и неподвижной, срастаясь с окружающими тканями. Подвижность теряет вместе с нею и кожа, её покрывающая. Подобного рода односторонние опухания подчелюстной железы наблюдаются при хроническом сапе с локализацией в одной какой-либо половине носа; кроме того, их иногда обнаруживают при хронических катарах верхнечелюстной пазухи, при односторонних катарах носа, распадающихся опухолях носа.
Двусторонние деформации лимфатических желез также развиваются в результате сапной инфекции при обоесторонних поражениях слизистой оболочки коса.
В некоторых случаях, в связи с изменением соответствующих лимфатических узлов, обнаруживают воспалительные инфильтрации связанных с ними лимфатических сосудов (лимфангиоит), которые, набухая, выступают в виде болезненных тяжей, прерывающихся узловатыми вздутиями, абсцессами, язвами. Покрывающая их кожа оказывается отёчною, малоподвижною и болезненною, а лимфатические узлы инфильтрированными.
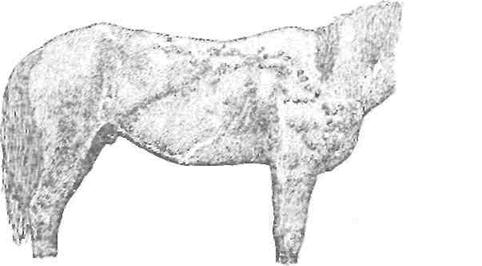
Рис. Язвы кожи при злокачественном лимфангиоите (по Velu).
Крайне резкие явления лимфангиоита обнаруживают при хроническом и остром сапе и африканском сапе лошадей, иногда при мыте, а ограниченные набухания на лице при пустулёзном стоматите, фолликулярном рините и на конечностях иногда при мокрецах.